Genetics!
•Télécharger en tant que PPT, PDF•
4 j'aime•4,109 vues

Genetics!!! It's all about the genetics!

Recommandé
Contenu connexe
Tendances
Tendances (12)
En vedette
Genetics!
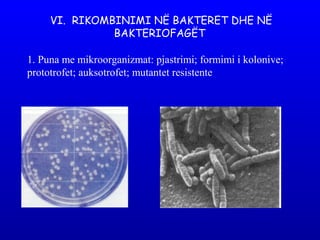
- 1. VI. RIKOMBINIMI NË BAKTERET DHE NË BAKTERIOFAGËT 1. Puna me mikroorganizmat: pjastrimi; formimi i kolonive; prototrofet; auksotrofet; mutantet resistente
- 2. Midis qelizave bakteriale mund të shkëmbehet material gjenetik nëpërmjet tre proçeseve kryesore: -Transformimi ( Shdrrimi): qelizat marrëse të materialit gjenetik që arrijnë të transformohen quhen transformantë - Konjugimi: qelizat marrëse të materialit gjenetik që arrijnë të ndryshohen quhen transkonjugantë - Transduktimi: qelizat marrëse të materialit gjenetik që arrijnë të ndryshohen quhen transduktantë
- 4. - Dukuria e transformimit përdoret për të përcaktuar lidhjen midis gjeneve në bazë të bashkëtransfor mimit.
- 5. Përckatimi i renditjes mbi bazën e shpeshtisë së bashkëtransformimit
- 6. a) F mund te dyfishoje ADN e tij b) Qelizat qe mbajne faktorin F prodhojne zgjatime (pilus) qe lejojne kontaktin me qeliza te tipit F- c) F+ mund te transmetojne nje kopje te faktorit F ne qeliza te tipit F- d) Qelizat F+ nuk mund te kontaktohen me qeliza te tjera F+ e) Rastesisht faktori F mund te lere citoplazmen dhe perfshihet ne kromozomen bakteriale Vetite dalluese te faktorit F: Faktori “seksual” F 2. Konjugimi Bakterial
- 8. - Transferimi i faktorit F
- 9. Transferimi i markatoreve i ndihmuar nga faktori F a b Rastesisht faktori F perfshihet ne kromozom duke formuar situaten Hfr. Kur F bashke me kromozomin transferohen ata transmetojne te gjithe gjenet ne qelizen pritese
- 10. Disa qeliza me faktorin F te perfshire mund te transferojne markatore ne qelizat pritese
- 11. Zbulimi i transferimit te gjeneve Bakterialë
- 12. Domosdoshmeria e kontaktit fizik midis qelizave te linjave te ndryshme bakteriale • Pas pjastrimit ne mjedis minimal te qelizave nga te dy anet e tubit nuk u gjet asnje koloni prototrofe. Kjo deshmon domosdoshmerine e kontaktit fizik midis dy linjave.
- 13. Zbulimi i faktorit te pjellorise F. • Percaktimi i drejtimit te transferimit te gjeneve a) Provat e Lederberg dhe Tatum treguan shmangie te rendesishme nga perpjestimi 1 : 1 ne "kryqezimin’’ A x B gje qe tregonte se gjenet ishin te lidhur. b) Provat e William Heyes 1953: Linja A: met- thr+ leu+ thi+ x Linja B: met+ thr- leu- thi- Rikombinantet fitoheshin vetem ne rastin kur me streptomicine trajtohej linja A dhe jo kur trajtohej linja B. Kjo tregonte se rikombinimi ndodhte ne linjen B, pra drejtimi i transferimit te materialit gjenetik behej nga A ne B.
- 14. Percaktimi i renditjes se gjeneve me ane te konjugimit te nderprere Harta e lidhjes se gjeneve te analizuar
- 15. Harta kromozomike rrethore e E.colit Faktori F mund të përfshihet në vende të ndryshme në gjenomën bakteriale: Linjat Hfr Gjenet që transferohen H origin thr pro lac pur gal his gly thi 1 origin thr thi gly his gal pur lac pro 2 origin pro thr thi gly his gal pur lac 3 origin pur lac pro thr thi gly his gal - Kjo mënyrë e transmetimit të gjeneve nga Hfr në F- mund të shpjegohet vetëm me faktin e gjendjes rrethore të kromozomit të Hfr:
- 16. Forma rrethore e kromozomës bakteriale
- 17. Kromozoma bakteriale eshte rrethore Te njejtet gjene mund te formojne linja te ndryshme Hfr te cilat dallohen vetem nga vendosja e faktorit F.
- 18. • Mekanizmi i mundshem i perfshirjes se faktorit F ne kromozomen bakteriale bartese te tij
- 19. Rikombinimi bakterial dhe hartezimi i kromozomit te E.coli • Rikombinimi midis gjeneve markatore pas transferimit • Gradienti i transferimit te gjeneve sherben per percaktimin e renditjes se gjeneve
- 20. Transmetimi i gjeneve markatore nga episomat • Rasti i seksduksionit ose transferimit te gjeneve nga faktori F
- 21. GJENETIKA E BAKTERIOFAGEVE • Infektimi i baktereve nga faget
- 22. Fagu i ngjitur me qelizen bakteriale Faget pasardhes brenda nje qelize bakteriale
- 24. Kryqezimi i fageve Tiparet ne studim: h - : mund te infektoje dy linja te ndryshme te E.coli (1 dhe 2) h+ : mund te infektoje vetem linjen 1 r- : lizis i shpejte pra formim i pllakave te medha r+: lizis i ngadalte, pra formim i pllakave te vogla • Nepermjet infektimit te dyfishte mund te analizohen gjenotipet ne faget pasardhes:
- 25. Kryqezimi i dyfishte ne faget Fenotipi Gjenotipi i mundshem Qarte, e vogel h- r+ Turbullt, e madhe h+ r- Turbullt, e vogel h+ r+ Qarte, e madhe h- r-
- 26. Lizogjenia • Bakteret lizogjenike • Situata profag • Faget virulente: plotesojne ciklin litik, ata mund te perfshihen ne kromozomen bakteriale ne formen e profagut duke i siguruar qelizes bartese rezistencen ndaj nje infektimi tjeter. • Faget e temperuar: plotesojne ciklin lizogjenik. Faget e temperuar mund te induktohen dhe te riaktivizohen
- 27. Cikle alternative te nje fagu ne qelizen bartese
- 28. Lidhja e fagut ne kromozomen bakteriale
- 29. 3.- TRANSDUKSIONI • Transduksioni i pergjithshem
- 31. NATYRA E GJENIT 1. Si punojne gjenet Cila eshte natyra e gjenit dhe si e kontrollojne gjenet fenotipin? Modeli yne per marredheniet midis gjenotipit dhe fenotipit: Tiparet karakteristike te nje organizmi percaktohen nga fenotipi i qelizave qe jane perberesit e pjeseve te tij. Fenotipi i qelizave percaktohet nga tere reaksionet kimike qelizore qe katalizohen nga enzimat. Funksioni i nje enzime percaktohet nga struktura e saj trepermasore e cila varet nga renditja e aminoacideve Enzimat dhe proteinat strukturore ne qelize percaktohen nga gjenotipi i qelizes Gjenet percaktojne renditjen e aminoacideve ne proteinat pra ata percaktojne fenotipin
- 32. Hipoteza Nje gjen nje enzime • Rruget biosintetike: • Kemi 5 mutante te paafte te sintetizojne produktin G : Mutantet Perberesit qe provohen A B C D E G 1 - - - + - + 2 - + - + - + 3 - - - - - + 4 - + + + - + 5 + + + + - +
- 33. a) Cila eshte renditja e perberesve ne kete vije biosintetike? Si mund ta zgjidhim kete problem? Tabela e mesiperme na sugjeron renditjen: E - A – C – B – D - G b) Ne cilen pike te vijes biosintetike eshte bllokuar secili nga mutantet e mesiperm? Tabela e mesiperme na sugjeron skemen e meposhtme: Eksperimentet e Beadle dhe Tatum Eksperimentet e Beadle dhe Tatum çuan ne perfundimin se: gjenet kontrollojne reaksionet biokimike duke kontrolluar prodhimin e enzimave
- 34. Skema e proves se Beadle dhe Tatum me Neurospora crassa
- 35. Mutacionet per argininen hartezonin ne tre vende te ndryshme ne kromozome te veçanta; po i quajme ata tre lokuse te ndryshme: arg-1, arg-2, arg-3 perkatesisht. Rritja e tyre realizohej sipas tabeles se meposhtme: Mutanti Shtesa Ornitine Citruline Arginine arg-1 + + + arg-2 - + + arg-3 - - +
- 36. Si rezultat i analizes mund te propozohet skema e meposhtme: Si perfundim mund te themi: • Reaksionet biokimike in vivo realizohen si nje seri reaksionesh te veçanta dhe te njepasnjeshme. • Çdo reaksion katalizohet ne menyre te veçante nga nje enzime e vetme. • Çdo enzime percaktohet nga nje gjen i veçante.
- 37. Gjenet dhe metabolizmi qelizor: semundjet gjenetike Pasojat e nderprerjes se disa rrugeve metabolike tek njeriu
- 38. Dukuri gjenetike qe shpjegohen sipas struktures se enzimave • Alelet e ndjeshme ndaj temperatures • Perpjestimet gjenetike: Shpjegimi i perpjestimit 9 : 7 • Dominanca dhe recesiviteti
- 39. Struktura e holle e gjenit Sipas teorise rruazore gjeni gezon vetite e meposhtme: • Eshte njesi themelore e struktures, e pandashme nga kryqkembimi. • Eshte njesi themelore e mutacionit, mund te ndryshoje nga nje forme alelike ne nje forme tjeter. • eshte njesi themelore e funksionit, pjese te gjenit, ne se egzistojne, nuk mund te funksionojne. Punimet e Saymor Benzerit me 1950 treguan se teoria e rruazave nuk ishte korrekte duke demostruar ndashmerine e metejshme te gjenit ne njesi me te vogla si dhe duke futur koncepte te reja
- 40. Mutantet rII te fagut T4 Midis pllakave te shkaktuara nga format normale te fagut T4 dallojme formen tipike mutante (brenda katrorit)
- 41. Linja e fagut T4 Linjat e E.colit B K (λ) rII E madhe, e rregullt Jo pllake rII + E vogel, e çrregullt E vogel, e çrregullt Formimi i pllakave nga kombinime te ndryshme te E.colit dhe linjave te fagut T4
- 42. Perzgjedhja e bakteriofageve ne kryqezime te ndryshme gjenetike
- 43. Per te njehsuar shpeshtine e rikombinimit numerohen pllakat e formuara ne linjen E.coli K (rikombinantet normale) si dhe pllakat e formuara ne linjen B ku rriten edhe mutantet edhe normalet. Rikombinantet shumezohen per 2, per aresye se nuk dallojme dot mutantet e dyfishte.
- 44. Rikombinimi intragjenik • Benzeri analizoi 8 lloje te ndryshme mutantesh rII duke i kryqezuar midis tyre ne te gjitha kombinimet dhe duke ndertuar harten e renditjes se tyre • Rikombinimi ndodhte brenda te njejtit gjen rII. Ne kete menyre kryqezimi midis nje mutanti a1 dhe nje tjetri a2 mund te simbolizohet: • Pra gjeni eshte i ndashem ne nen njesi me ane te rikombinimit. Shpeshtia me e vogel e rikombinimit midis nje cifti mutantesh alele ishte 0,01%.
- 45. Vendet mutacionale Perdorimi i delecioneve ne hartezimin e vendeve mutacionale • Analizojme disa gjene qe renditen keshtu ne harten gjenetike • Kur nje mutant D1 nuk jep rikombinante rII+ me mutantet qe kane vendet mutacionale 1,2,3,4,5,6,7,8 ai ka nje delecion ne zonen 1 - 8 • Kur një mutant D2 nuk jep rikombinante rII+ me mutantet 5,6,7,8,89,10,11, 12 ai ka delecion ne zonen 5 - 12
- 46. Duke i mbivendosur dy delecionet e mesiperme mund te percaktojme zonat e meposhtme te gjenit. • Nje mutant i ri qe jep rikombinante rII+ kur kryqezohet me D1 por jo me D2 duhet te kete nje mutacion ne zonen III. • Nje mutant i ri qe jep rikombinante rII+ me D2 por jo me D1 duhet te kete nje mutacion ne zonen I. • Nje mutant i ri qe nuk jep rikombinante rII+ as me D1 as me D2 duket te kete nje mutacion ne zonen II.
- 47. Pas percaktimit te zones se nje mutacioni ai mund te hartezohet ne lidhje me mutacionet e tjera duke formuar nje harte te hollesishme
- 48. Analiza e vendeve mutacionale Shperndarja e 1612 mutacioneve te rastesishme ne lokusin rII , çdo ngjarje mutacionale pasqyrohet me ane te nje katrori te vogel
- 49. Struktura e holle e lokusit rII (permbledhje)
- 51. Skema e perplotesimit ne mutantet e fagut T4
- 52. Skema qe shpjegon perplotesimin ne lokusin rII
- 54. Cistroni Cistroni eshte nje vend gjenetik ne te cilin dy mutacione ne pozicionin trans nuk mund te perplotesohen.
- 55. ADN-ja RIKOMBINANTE Enzimat e restriksionit Enzimat e restriksionit presin ne vende me renditje te veçanta duke prodhuar nje popullate fragmentesh me funde identike
- 56. Si pritet ADN-ja në copa - Enzimat e restriksionit, roli dhe përdorimi i tyre në inxhinierinë gjenetike - Palindromat dhe veprimi i enzimave të restriksionit
- 58. Tipi Aktiviteti i enzimës Kërkesë për ATP Vendi i prerjes I Prerje dhe metilim Po Rastësisht larg nga vendi i njohjes II Vetëm prerje Jo Brenda vendit të njohjes III Prerje dhe metilim Po Rastësisht pranë vendit të njohjes - Tipe të ndryshëm të enzimave të restriksionit
- 59. Prerja dhe ngjitja e copave të ADN nga enzimat e ndryshme
- 60. Prerje e ADN-së gjatësore të hapur - Numri i vendeve të prerjes është i lidhur me numrin e copave të prodhuara kur ADN-ja pritet nga një enzimë restriksioni
- 62. Vektoret Vetite e vektoreve: • nje molekule e vogel, e karakterizuar mire qe permban nje origjine dyfishimi e cila lejon vetdyfishimin duke perfshire dhe fragmentet e huaja qe ajo permban • lejon te fitohet lehtesisht nje molekule hibride Plazmidet Fagu λ Kozmidet (λ + plazmid) Faget me nje varg ADN-je
- 63. Vektori klonues Madhësia e ADN që klonohet Metoda e shtimit Futja në qelizat bakteriale Plazmid Deri 15 kb Dyfishim i plazmidit Transformim Fagu lambda Deri 23 kb Riprodhim i fagut Infektim me anë të fagut Kozmid Deri 44 kb Riprodhim i plazmidit Infektim me anë të fagut - Disa të dhëna mbi vektorët
- 64. Metodologjia e ADN-se rikombinante Prodhimi i librarise gjenike dhe izolimi i klonit te desheruar
- 66. Zbulimi i gjeneve te klonuar (Southern blotting)
- 67. Teknika te tjera: cADN Northern blotting, Western blotting,
- 68. Teknika Maxam-Gilbert e sekuencimit
- 70. Sekuencimi sipas metodës dideoksi të Sanger - Pergatitja e ADN stampë që do të sekuencohet
- 71. - Reaksioni i zgjatjes së vargut dhe analiza në xhel
- 72. Shumefishimi i gjeneve, teknika PCR (polymerase chain reaction)
- 73. Zbatime te teknologjise se ADN-se rikombinante ne eukariotet • Vektoret e majase
- 74. Kafshet transgjenike: nje shembull i terapise gjenike tek minjte Shprehja e gjenit per hormonin e rritjes pas futjes me ane te plazmidit
- 75. Kerkimi per semundje te ndryshme Amniocenteza
- 76. Ndryshimi i vendeve te restriksionit Rasti i anemise falciforme
- 77. Perdorimi i RFLP
- 78. ADN FINGERPRINTS - Përdorimi i mikrosatelitëve - Llojet ndryshme të RFLP-ve mund të analizohen me anë të prerjes me enzima dhe vrojtimit të brezave elektroforetikë. - Per çdo individ formohet një model i veçantë brezash
- 82. Gjenomika: shprehja e gjeneve dhe mikroarray - mjetet e rinj në dispozicion si: sekuenciatorët e ADN, microarray të ADN ose chip janë në gjendje të analizojnë shpejt dhe me saktësi mijëra gjene. - çfarë është dhe si funksionon chip-i i ADN-së?
- 83. Skema e një çipi
- 84. Analiza e proçesit të ibridizimit identifikon gjenet që përgjigjen në mënyrë të veçantë
- 85. Analiza e kompjukterizuar e renditjeve lejon identifikimin e gjeneve që i përgjigjen një trajtimi me lëndë të ndryshyme
- 86. GJENET NË POPULLATA Popullata si një rezervuar gjenetik Llojet e ndryshme janë të ndarë në grupe ose popullata (deme). Popullatat janë dinamike. Popullatat mund të përshkruhen në terma të dëndësisë, shpërndarjes hapsinore, ritmeve të lindjeve dhe vdekjeve si dhe nëpërmjet tipareve gjenetike
- 87. Popullata, natyrisht ka më shumë informacion se individi. Koncepti i fondit gjenetik: kompleti i informacionit gjenetik që mban një popullatë e caktuar. Çdo popullatë mund të ndryshojë si rezultat i kushteve që çojnë në ndryshimin e shpeshtive alelike. Si mund ta karakterizojmë një popullatë në terma gjenetikë?
- 88. Raste të ndryshme të njehsimit të: Shpeshtive fenotipike Shpeshtive gjenotipike Shpeshtive alelike
- 89. Ligji i Hardy – Weinberg-ut Është një model matematik që siguron mënyrat e vlerësimit të shpeshtive alelike në popullatë dhe përshkruan si kombinohen alelet për të formuar gjenotipet. Ky model bazohet në këto kushte: - Popullata me përmasa të mëdha - Mungon seleksioni - Kryqëzimet janë të rastit - Mungojnë mutacionet - Mungojnë migrimet
- 90. Shembull: Analizohet një gjen me dy alele: A dhe a (dominanca e plotë). Shënojmë përkatësisht me p dhe q shpeshtitë e aleleve A dhe a. Meqenëse kemi vetëm dy alele është i vërtetë rregulli: p (A) + q (a) = 1 p (A) Vezët q (a) Spermatozoidët p (A) q (a) AA p2 Aa pq Aa pq Aa q2
- 91. p2 tregon shpeshtinë e homozigotëve dominantë (AA) 2pq tregon shpeshtinë e heterozigotëve (Aa) q2 tregon shpeshtinë e homozigotëve reçesivë (aa) Në mënyrë të përmbledhur shpërndarjen e gjenotipeve të ndryshëm mund të shkruajmë: p2 + 2pq + q2 = 1 AA Aa aa
- 92. Shembull numerik: duke u nisur nga shpeshtitë fillestare p=0.6 dhe q= 0.4 në brezninë pasardhëse shpeshtitë gjenotipike do të jenë: p (A)= 0.6 Vezët q (a)= 0.4 Spermatozoidët p (A) = 0.6 q (a) = 0.4 AA p2 = 0.36 Aa pq = 0.24 Aa pq = 0.24 Aa q2 = 0.16
- 93. Shpeshtitë alelike në këtë brezni janë: p(A) = p2 + 1/2 (2pq) = 0.36 + 1/2 (0.48) = 0.6 q(a) = q2 + 1/2 (2pq) = 0.16 + 1/2 (0.48) = 0.4 Sidoqoftë shpeshtitë gjenotipike edhe në këtë brezni janë po ato të breznisë paraardhëse.
- 94. Përdorimi i ligjit të Hardy –Weinberg-ut në Gjenetikën Humane Alelet autosomike kodominante (sistemi MN) Alelet autosomike dominante dhe reçesive (fibroza kistike: aa=1/2500) Tiparet e lidhur me seksin (daltonizmi: Xa = 0.08) Alelet e shumfishta (sistemi AB0)