

Recommandé
Contenu connexe
Tendances
Tendances (20)
Similaire à Membrana Plasmática
Similaire à Membrana Plasmática (17)
Membrana Plasmática
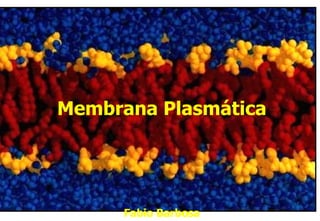
- 1. Membrana Plasmática Fabio Barbosa
- 2. Membrana Celular A principal função da membrana celular é manter, de forma seletiva, moléculas tão diversas como proteínas e pequenos solutos, no interior da célula. Assim, a membrana funciona de forma eficiente para regular seletivamente sua permeabilidade. A composição da membrana celular tem sido estudada de forma intensa, a partir do uso de diversas técnicas físicas e químicas, discutiremos a seguir a evolução dos principais modelos da membrana celular. CHE
- 3. Membrana Celular No livro clássico de Oparin, A Origem da Vida, esse propôs que para qualquer forma de vida, das mais simples que sejam, faz-se necessária uma barreira física, que separe a parte viva do meio que a cerca. Esse trabalho destaca a necessidade de uma membrana para isolar, até mesmo as formas de vida mais simples, do meio exterior. CHE
- 5. MEMBRANA PLASMÁTICA Composição química LIPÍDIOS, PROTEÍNAS E AÇÚCARES
- 6. Modelos de Membrana Celular Modelo de Robertson (1957). O estudo de eritrócitos realizados por Gorter & Grendel em 1925 indicou que o conteúdo lipídico das membranas ocupava uma área duas vezes maior que a superfície da célula. Tal observação levou à hipótese da bicamada lipídica, com a parte polar voltada para os meios intra e extra celular e a parte hidrofóbica voltada para o interior da membrana, escondida do solvente. Posteriormente Schmitt e colaboradores, a partir de estudos de polarização da luz, propuseram que eritrócitos apresentavam lipídios perpendiculares ao plano da membrana, como espera-se de uma bicamada (Schmitt et al ., 1937, 1938). Outros cientistas propuseram a presença de proteínas nas membranas (Danielli & Davson, 1935), com a participação protéica estendendo-se até 60 % da membrana. Baseado nessas informações Robertson (1957, 1981) propôs que as proteínas estivessem distribuídas sobre a superfície da membrana. CHE
- 7. Modelos de Membrana Celular O modelo de Robertson era coerente com a informação sobre a presença de proteínas nas membranas, bem como com a presença da bicamada lipídica, contudo falhava ao colocar proteínas globulares na superfície da membrana. A presença de uma camada de proteína na membrana formava uma blindagem na superfície da membrana, o que impossibilita a comunicação entre os meios intra e extra-celular. Referências: Robertson , J. D. (1957). J. Biophys . Biochem . Cytol ., 3:1043-1048 . Robertson , J. D. (1981). J. Cell Biol . 91(3 Pt 2):189s-204s. Review. Proteína globular Bicamada lipídica CHE
- 8. Modelos de Membrana Celular Modelo de Stein & Danielli. Esse modelo propõe a presença de um canal transmembrana composto por proteínas, o que permite comunicação entre os meios intra e extra celular, aumentando a permeabilidade da membrana celular. Nesse modelo toda a membrana é revestida por proteínas Stein & Danielli (1956). O principal problema desse modelo é o envolvimento de toda a membrana celular por proteínas, não há contato para porção polar da bicamada lípídica com o solvente do meio extracelular, ou com o citoplasma, a bicamada lipídica fica blindada pela proteína. Referência: Stein, W. D. & Danielli, J. F. (1956). Discuss. Faraday Soc . 21:238-251. Fonte: Garcia, E. A. C. Biofísica. Editora Savier, 2000 (pg. 5). CHE
- 9. Modelo de Lucy & Glauert. Nesse modelo o lipídeos formam micelas globulares, que estão revestidas por proteínas ( Lucy & Glauert , 1964 ). Esse modelo apresenta o mesmo problema do modelo de Robertson, não há comunicação entre os meios extra e intra celular. Referências: Lucy , J. A. & Glauert , A. M. (1964). J. Mol. Biol . 12: 727-748. De Mello, W. C. (1972). Electrical phenomena in the heart, Academic Press, New York and London. Fonte: Garcia, E. A. C. Biofísica. Editora Savier, 2000 (pg. 5). Modelos de Membrana Celular CHE
- 10. Fonte: Garcia, E. A. C. Biofísica. Editora Savier, 2000 (pg. 5). Modelo de Benson. Nesse modelo a membrana é composta por uma matriz protéica com lipídios dispersos. O principal problema dessa proposta está na inexistência de canais transmembranas. Referência: Benson , A. A. (1966). J. Amer . Oil Chem . Soc ., 43:265-270. Modelos de Membrana Celular CHE
- 11. Modelo de Lenard&Singer, , indicando duas proteína inseridas na bicamada lipídica (elipsóides cinzas). A proteína da esquerda é uma proteína extrínseca e a da direira uma proteína intrínseca. Os fosfolipídios são indicados com a cabeça polar em preto e a cauda hidrofófica pelas linhas que saem da esfera preta. Modelo de Lenard & Singer. Esse modelo, mais realístico, prevê a presença de proteínas numa bicamada lipídica (Lenard & Singer, 1966), há previsão de proteínas transmembranas que atravessam a bicamada lipídica. Referência: Lenard, J. & Singer, S. J. (1966). Proc. Natl. Acad. Sc. 56:1828-1835. Modelos de Membrana Celular CHE
- 12. Referência: Singer, S. J. & Nicolson , G. L. (1972) Science , 175:720-731. Proteína intrínseca, ou transmembrana Proteínas extrínsecas Modelo de mosaico fluido. Experimentos mais detalhados mostraram deficiências nos diversos modelos de membrana celular. Singer e Nicolson (1972) propuseram um modelo de membrana constituído de uma bicamada lipídica, onde encontram-se inseridas proteínas. Há dois tipos de proteínas inseridas na membrana, uma que atravessa toda a membrana, chamada proteína intrínseca, ou transmembrana. O segundo tipo de proteína localiza-se sobre a membrana, sendo encontrada tanto no exterior como voltada para o citoplasma. Esse segundo tipo de proteína é chamado extrínseca. Modelos de Membrana Celular CHE
- 13. Referência: Singer, S. J. & Nicolson , G. L. (1972) Science , 175:720-731. Proteína intrínseca, ou transmembrana Proteínas extrínsecas Esse modelo prevê a passagen seletiva de íons pelas proteínas intrínsecas, que são chamadas de canais ou bombas como veremos em detalhe no estudo do potencial de membrana. Outra característica desse modelo é liberdade de movimentação das proteína na bicamada. De acordo com características básicas do modelo, mosaicismo e difusão, previu-se a liberdade lateral e rotatória, assim como a distribuição aleatória de componentes moleculares na membrana. Modelos de Membrana Celular CHE
- 14. O modelo de mosaico fluido é usado até hoje, apesar de experimentos posteriores, terem deixado claro que a liberdade de movimentação das proteínas transmembranas e extrínsecas, não representa a realidade observada ( Vereb et al., 2004 ). Uma das características da membrana celular, não previstas no modelo de mosaico fluido, é que as proteínas apresentam uma distribuição não aleatória, há uma concentração de proteínas em regiões definidas na membrana, conhecidas como balsas lipídicas. Essas balsas lipídicas são pequenas da membrana, onde alguns lipídios (normalmente esfingolipídeos e colesterol) e proteínas estão concentrados. Modelos de Membrana Celular Balsas lipídicas, mostrando setores da membrana que fixam essas nas regiões indicas. Fonte: Vereb et al. 2004 . CHE
- 15. Modelo Atual da Membrana Celular Um modelo para membrana celular mais realístico prevê além da clássica bicamada lipídica e as proteínas transmembranas e extrínsecas os seguintes aspectos: 1) Distribuição não aleatória das proteína na membrana. O modelo original de mosaico fluido prevê uma distribuição aleatórias das proteína na bicamada lipídica. 2) Contatos moleculares quase permanentes. 3) Domínios de membrana: difusão limitada, reorganização dinâmica. 4) Plataformas lipídicas. 5) Proteínas são importantes elementos estruturais. 6) Estruturação dinâmica. CHE
- 16. Proteína da classe Cadeia lateral de oligossacarídeo Glicolipídio Fosfolipídeo Proteína globular Segmento hidrofóbico da proteína Colesterol Modelo Atual da Membrana Celular CHE
- 17. Modelo de Singer e Nicolson (1972) Modelo do Mosaico Fluido Proteínas embebidas na bicamada lipídica; Estrutura molecular da membrana plasmática CHE
- 18. CHE Proteínas Integrais (transmembranas) Periféricas Lipídeos Glicolipídeos Colesterol Fosfolipídeos Fosfatidilcolina Fosfatidiletanolamina Fosfatidilserina Esfingomielina Proteína / Lipídeo • Proporção variável
- 19. Composição Lipídica de algumas membranas celulares
- 20. Hidrofílica (cabeça) Hidrofóbica (caudas) CHE Moléculas Anfipáticas LIPÍDEOS DE MEMBRANAS
- 21. Fosfolipídeos CHE Fosfatidilcolina Fosfatidiletanolamina Fosfatidilserina Esfingomielina
- 22. Flip Flop Rotação Difusão Lateral dependente da temperatura 1- Fluidez da membrana Fluido Bidimensional movimentação dos fosfolipídeos dentro da bicamada
- 23. Composição Fosfolipídica Natureza das caudas de hidrocarbonetos Caudas curtas (maior fluidez) que caudas longas Insaturação (maior fluidez) que saturação INSATURADOS - viscosa + fluida SATURADOS + viscosa - fluida 1- Fluidez da membrana CHE
- 24. 1- Fluidez da membrana Colesterol Modula a fluidez das membranas em células animais Enrijece a bicamada lipídica, tornando-a menos fluida e menos permeável
- 25. Diferenças na composição da bicamada entre as faces citosólica e extracelular 2- Assimetria da Bicamada Lipídica
- 26. Barreira hidrofóbica impermeável a solutos e íons tamanho da molécula solubilidade da molécula (em óleo) 3- Permeabilidade da Bicamada Lipídica CHE
- 28. Proteínas de membrana K + Na +
- 29. Proteínas Transmembrana Moléculas anfipáticas ligadas covalentemente aos lipídeos Proteínas -Hélice
- 30. Proteínas Transmembrana Proteínas receptoras : cruza a membrana uma única vez Proteínas Transmembrana
- 31. Poro Hidrofílico : múltiplas -Hélices formam poros aquosos Proteínas Transmembrana
- 32. Movimentação das proteínas na bicamada Propriedades das Proteínas de membrana 1- Mobilidade
- 33. Restrição de movimento das proteínas, confinando-as em locais específicos 2- Domínios de membrana
- 35. Hidratos de carbono ligados covalentemente aos lipídeos e proteínas Glicoproteínas Glicolipídeos Proteoglicanas oligossacarídeos polissacarídeos glicosaminoglicanas GLICOCÁLICE OU GLICOCÁLIX Açucares de Membrana
- 36. GLICOCÁLICE
- 37. Funções do Glicocálice - proteção e lubrificação da superfície celular - reconhecimento célula-célula e adesão celular
- 38. - propriedades enzimáticas (peptidase/glicosidase) - especificidade do sistema sanguíneo ABO; - alteração da superfície em células cancerígenas; - ligação de toxinas, vírus e bactérias; Funções do Glicocálice
- 39. Interação de Peptídeos Tóxicos com a Membrana Celular A membrana celular animal apresenta uma bicamada lipídica de aproximadamente 60 Å de extensão, o que possibilita que proteínas como o centro de reação fotossintético atravessem a membrana, contudo peptídeos pequenos, como os mastoparanos e o anoplin, possuem comprimento de 21 e 15 Å, respectivamente, não permitindo que esses peptídeos atravessem a membrana celular. Resta a questão sobre a forma de ação desses peptídeos, visto que evidências experimentais indicam que os mesmos atuem na membrana celular, desestabilizando-a. Uma possível forma de ação desses peptídeos, é por meio do desmonte da camada externa da membrana, o que levaria à sua desestruturação e consequente quebra da membrana celular. A forma exata da ação dos peptídeos tóxicos com a membrana celular é ainda matéria de debate. Um possível modelo de interação do peptídeo com a bicamada está mostrado nos slides seguintes.
- 40. ~60 Å Ancoragem do peptídeo na membrana celular. Interação de Peptídeos Tóxicos com a Membrana Celular
- 41. ~60 Å Início da desestabilização da membrana celular. Interação de Peptídeos Tóxicos com a Membrana Celular
- 42. ~60 Å Início da desmontagem da primeira camada lipídica. Interação de Peptídeos Tóxicos com a Membrana Celular
- 43. ~60 Å Início da desmontagem da segunda camada lipídica. Interação de Peptídeos Tóxicos com a Membrana Celular
- 44. ~60 Å Desmontagem da bicamada lipídica. Interação de Peptídeos Tóxicos com a Membrana Celular
- 45. ~60 Å Início da passagem de citoplasma para o meio exterior e entrada de substâncias do meio extra-celular. Interação de Peptídeos Tóxicos com a Membrana Celular
- 46. ~60 Å Abertura de um grande canal transmembrana. Interação de Peptídeos Tóxicos com a Membrana Celular
- 47. ~60 Å Fluxo de substâncias para o interior e exterior da célula leva à apoptose. Interação de Peptídeos Tóxicos com a Membrana Celular
- 48. Streptolisina A toxina streptolisina é uma proteína produzida pelo Streptococcus pyrogenes de massa molecular 65 kDa. Essa toxina liga-se à membrana celular, que apresenta colesterol na sua superfície. Depois de ligar-se à membrana a streptolisina polimeriza-se, formando poros na membrana celular (figura ao lado). Esses poros transmembranas podem ter até 350 Å de diâmetro. Os diagramas esquemáticos nos próximos slides ilustram os principais passos na formação dos poros. Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. a) Membrana de eritrócito lisadas por streptolisina. b) Oligômeros de streptolisina, mostrando diversas estruturas de bastões. a) b)
- 49. Processo de formação de poros pela streptolisina. 1) Associação reversível das streptolisinas monoméricas com membranas celulares contendo colesterol. 1 Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. Streptolisina
- 50. Processo de formação de poros pela streptolisina. 2) Dois monômeros reagem formando um dímero inserido na membrana. 1 2 Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. Streptolisina
- 51. Processo de formação de poros pela streptolisina. 3) Oligomerização segue pela adição sucessiva de monômeros. 1 2 3 Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. Streptolisina
- 52. Processo de formação de poros pela streptolisina. 4) Há formação de um poro transmembrana. 1 2 3 4 Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. Streptolisina
- 53. Representação esquemática do dano causado na membrana pela ação do oligômero de streptolisina. Referência: Bhakdi , S., Tranum-Jensen , J., & Sziegoleit , A. (1985). Infec . Immun . 47:52-60. Streptolisina
- 54. Membrana Plasmática: Especializações de membrana
- 55. ESPECIALIZAÇÕES DA MEMBRANA PLASMÁTICA SUPERFÍCIE APICAL DA CÉLULA SUPERFÍCIE BASO-LATERAL DA CÉLULA 1- Microvilosidades 2- Cílios/Flagelos 3- Estereocílios 1- Junções celulares Junções célula-célula Junções célula-matriz extracelular
- 56. ESPECIALIZAÇÕES DA SUPERFÍCIE APICAL DA MEMBRANA MICROVILOSIDADES -Projeções cilíndricas do citoplasma, envolvidas por membrana que se projetam da superfície apical da célula -São imóveis -Aumentam a área de superfície celular -Filamentos de actina
- 58. ESPECIALIZAÇÕES DA SUPERFÍCIE APICAL DA MEMBRANA ESTEREOCÍLIOS -São parecidos com microvilosidades- mais longas e ramificadas -São imóveis -Encontrados no epidídimo e nas células pilosas do ouvido interno -Aumentam a área de superfície das células -Filamentos de actina mais discretos que nas microvilosidades
- 59. CÍLIOS/FLAGELOS
- 60. ESPECIALIZAÇÕES DA SUPERFÍCIE APICAL DA MEMBRANA CÍLIOS -Projeções cilíndricas MÓVEIS, semelhantes a pêlos -Função: propulsão de muco e de outras substâncias sobre a superfície do epitélio, através de rápidas oscilações rítmicas e no caso dos flagelos funcionam na locomoção -Microtúbulos organizados (9 + 2), inseridos no corpúsculo basal
- 61. ESPECIALIZAÇÕES DA SUPERFÍCIE BASO-LATERAL DA CÉLULA JUNÇÕES CELULARES JUNÇÃO OCLUSIVA JUNÇÕES JUNÇÃO ADERENTE DESMOSSOMA JUNÇÃO COMUNICANTE COMPLEXO JUNCIONAL Matriz extracelular
- 62. Une as células formando uma barreira impermeável JUNÇÃO OCLUSIVA Evita movimentação de moléculas entre diferentes domínios de membrana
- 63. Cinturão de adesão apical, abaixo junção oclusiva JUNÇÃO ADERENTE JUNÇÕES CELULARES ADESÃO
- 64. Placas de adesão em forma de disco DESMOSSOMAS JUNÇÕES CELULARES ADESÃO
- 65. JUNÇÃO COMUNICANTE * Formada por 6 proteínas transmembranas – conexinas * Regulada abrem e fecham