

Recommandé
Contenu connexe
Tendances
Tendances (20)
En vedette
En vedette (20)
Similaire à Auditory receptors
Similaire à Auditory receptors (20)
Plus de DinDin Horneja
Plus de DinDin Horneja (20)
Dernier
Dernier (20)
Auditory receptors
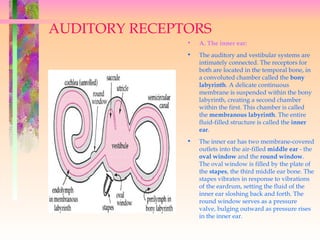
- 1. AUDITORY RECEPTORS • A. The inner ear: • The auditory and vestibular systems are intimately connected. The receptors for both are located in the temporal bone, in a convoluted chamber called the bony labyrinth. A delicate continuous membrane is suspended within the bony labyrinth, creating a second chamber within the first. This chamber is called the membranous labyrinth. The entire fluid-filled structure is called the inner ear. • The inner ear has two membrane-covered outlets into the air-filled middle ear - the oval window and the round window. The oval window is filled by the plate of the stapes, the third middle ear bone. The stapes vibrates in response to vibrations of the eardrum, setting the fluid of the inner ear sloshing back and forth. The round window serves as a pressure valve, bulging outward as pressure rises in the inner ear.
- 2. The oval window opens into a large central area within the inner ear called the vestibule. All of the inner ear organs branch off from this central chamber. On one side is the cochlea, on the other the semicircular canals. The utricle and saccule, additional vestibular organs, are adjacent to the vestibule. The membranous labyrinth is filled with a special fluid called endolymph. Endolymph is very similar to intracellular fluid: it is high in potassium and low in sodium. The ionic composition is necessary for vestibular and auditory hair cells to function optimally. The space between the membranous and bony labyrinths is filled with perilymph, which is very much like normal cerebral spinal fluid. B. Auditory transduction: The transduction of sound into a neural signal occurs in the cochlea. If we were to unroll the snail-shaped cochlea, it would look like this: As the stapes vibrates the oval window, the perilymph sloshes back and forth, vibrating the round window in a complementary rhythm. The membranous labyrinth is caught between the two, and bounces up and down with all this sloshing. Now let's take a closer look at the membranous labyrinth. If we cut the cochlea in cross section, it looks like this:
- 3. The membranous labyrinth of the cochlea encloses the endolymph-filled scala media. The two compartments of the bony labyrinth, which house the perilymph, are called the scalae vestibuli and tympani. Within the scala media is the receptor organ, the organ of Corti. It rests on part of the membranous labyrinth, the basilar membrane. A single turn of the cochlea has been outlined in blue. You can see the auditory nerve exiting at the base of the cochlea; it will travel through the temporal bone to the brainstem. The auditory hair cells sit within the organ of Corti. There are inner hair cells, which are the auditory receptors, and outer hair cells, which help to "tune" the cochlea, as well as supporting cells. The sensitive stereocilia of the inner hair cells are embedded in a membrane called the tectorial membrane. As the basilar membrane bounces up and down, the fine stereocilia are sheared back and forth under the tectorial membrane. When the stereocilia are pulled in the right direction, the hair cell depolarizes. This signal is transmitted to a nerve process lying under the organ of Corti. This neuron transmits the signal back along the auditory nerve to the brainstem. As with almost all sensory neurons (the exception is in the retina), its cell body lies outside the CNS in a ganglion. In this case, the ganglion is stretched out along the spiralling center axis of the cochlea, and is named the spiral ganglion. You can see most of the structures in this higher magnification of the organ of Corti; unfortunately, the inner hair cells have been artifactually pulled away from the tectorial membrane.
- 4. The basilar membrane is actually thinner and narrower at the base of the cochlea than at the tip (apex), which seems backwards given that the cochlea is widest at the base. The properties of the basilar membrane change as its shape changes; just as with guitar strings, thin things vibrate to high pitches, and thick things vibrate to low pitches. This means that the basilar membrane vibrates to high frequencies at the base of the cochlea and to low frequencies at the apex. A hair cell at the base of the cochlea will respond best to high frequencies, since at those frequencies the basilar membrane underneath it will vibrate the most. The key idea is that although the hair cells are arranged in order along the basilar membrane, from high-frequency to low-frequency, it is the properties of the basilar membrane that set up this gradient, not the properties of the hair cells. Our ability to discriminate two close frequencies is actually much better than one would predict just from the mechanics of the basilar membrane. One theory to explain the mystery is that the outer hair cells help to "sharpen the tuning". Outer hair cells can actually move (change length) in response to nerve stimulation. If they could push the basilar membrane up and down, they could amplify or damp vibrations at will, making the inner hair cells more or less responsive. (Just like you can push a child higher and higher on a swing or bring her to a halt - it's all in when you push.) An interesting philosophical question here is, if the outer hair cells can move the basilar membrane, can that in turn move the oval window? And the stapes? And the eardrum? Can the ear, in fact, work in reverse and become a speaker? You may laugh, but there has been at least one case in the history of medicine of a patient complaining of persistent whispering in her ear. She was dismissed as crazy, until one obliging doctor finally put his stethoscope to her ear and listened. He could hear the whispering too. You can draw your own moral from this story. However, most cases of tinnitus (a persistent ringing, whistling, or roaring in the ears) are not audible to the examiner. Little is known about the phenomenon, which is unfortunate because it can be very distressing to the sufferer.
- 5. C. Central auditory pathways: The auditory nerve carries the signal into the brainstem and synapses in the cochlear nucleus. From the cochlear nucleus, auditory information is split into at least two streams, much like the visual pathways are split into motion and form processing. Auditory nerve fibers going to the ventral cochlear nucleus synapse on their target cells with giant, hand-like terminals. Something about this tight connection allows the timing of the signal to be preserved to the microsecond (action potentials are on the order of milliseconds, so it is no mean feat). The ventral cochlear nucleus cells then project to a collection of nuclei in the medulla called the superior olive. In the superior olive, the minute differences in the timing and loudness of the sound in each ear are compared, and from this you can determine the direction the sound came from. The superior olive then projects up to the inferior colliculus via a fiber tract called the lateral lemniscus. The second stream of information starts in the dorsal cochlear nucleus. Unlike the exquisitely time-sensitive localization pathway, this stream analyzes the quality of sound. The dorsal cochlear nucleus, with fairly complex circuitry, picks apart the tiny frequency differences which make "bet" sound different from "bat" and "debt". This pathway projects directly to the inferior colliculus, also via the lateral lemniscus.
- 6. Notice that both pathways are bilateral. The consequence of this is that lesions anywhere along the pathway usually have no obvious effect on hearing. Deafness is essentially only caused by damage to the middle ear, cochlea, or auditory nerve. From the inferior colliculus, both streams of information proceed to sensory thalamus. The auditory nucleus of thalamus is the medial geniculate nucleus. The medial geniculate projects to primary auditory cortex, located on Keep in mind, as you try to remember this pathway, that the auditory nuclei all seem to have counterparts in the banks of the temporal lobes. other systems, making life confusing. Fibers from the cochlear nuclei and the superior olive (not the inferior) travel up the lateral lemniscus (not the medial) to the inferior colliculus (not the superior), and then to the medial geniculate (not the lateral). Try remembering the mnemonic, "S-L-I-M" . D. The vestibular system The purpose of the vestibular system is to keep tabs on the position and motion of your head in space. There are really two components to monitoring motion, however. You must be able to detect rotation, such as what happens when you shake or nod your head. In physics, this is called angular acceleration. You must also be able to detect motion along a line - such as what happens when the elevator drops beneath you, or on a more subtle note, what happens when your body begins to lean to one side. This is called linear acceleration. The vestibular system is divided into two receptor organs to accomplish these tasks. 1. The semicircular canals detect angular acceleration. There are 3 canals, corresponding to the three dimensions in which you move, so that each canal detects motion in a single plane. Each canal is set up as shown below, as a continuous endolymph-filled hoop. The actual hair cells sit in a small swelling at the base called the ampula. 2. The VOR Although the VOR works on all three muscle pairs, the medial-lateral rectus pair, coupled to the horizontal canal, is geometrically the easiest to draw
- 7. The Outer Ear Sound energy spreads out from its sources. For a point source of sound, it spreads out according to the inverse square law. For a given sound intensity, a larger ear captures more of the wave and hence more sound energy. The outer ear structures act as part of the ear's preamplifier to enhance the sensitivity of hearing. The auditory canal acts as a closed tube resonator, enhancing sounds in the range 2-5 kiloHertz. The Tympanic Membrane The tympanic membrane or "eardrum" receives vibrations traveling up the auditory canal and transfers them through the tiny ossicles to the oval window, the port into the inner ear. The eardrum is some fifteen times larger than the oval window, giving an amplification of about fifteen compared to the oval window alone.
- 8. Auditory Canal Resonance The maximum sensitivity regions of human hearing can be modeled as closed tube resonances of the auditory canal. The observed peak at about 3700 Hz at body temperature corresponds to a tube length of 2.4 cm. The higher frequency sensitivity peak is at about 13 kHz which is somewhat above the calculated 3rd harmonic of a closed cylinder The Ossicles The three tiniest bones in the body form the coupling between the vibration of the eardrum and the forces exerted on the oval window of the inner ear. With a long enough lever, you can lift a big rock with a small applied force on the other end of the lever. The amplification of force can be changed by shifting the pivot point. The ossicles can be thought of as a compound lever which achieves a multiplication of force. This lever action is thought to achieve an amplification by a factor of about three under optimum conditions, but can be adjusted by muscle action to actually attenuate the sound signal for protection against loud sounds.
- 9. Ossicle Vibration The vibration of the eardrum is transmitted to the oval window of the inner ear by means of the ossicles, which achieve an amplification by lever action. The lever is adjustable under muscle action and may actually attenuate loud sounds for protection of the ear. Inner Ear
- 10. The inner ear can be thought of as two organs: the semicircular canals which serve as the body's balance organ and the cochlea which serves as the body's microphone, converting sound pressure impulses from the outer ear into electrical impulses which are passed on to the brain via the auditory nerve. The basilar membrane of the inner ear plays a critical role in the perception of pitch according to the place theory. Semicircular canals The semicircular canals are the body's balance organs, detecting acceleration in the three perpendicular planes. These accelerometers make use of hair cells similar to those on the organ of Corti, but these hair cells detect movements of the fluid in the canals caused by angular acceleration about an axis perpendicular to the plane of the canal. Tiny floating particles aid the process of stimulating the hair cells as they move with the fluid. The canals are connected to the auditory nerve.
- 11. Sonar detection in bats HOMING IN. The distance between pressure pulses in a bat's ultrasonic chirps and echoes (represented by white and red wave outlines, respectively) determines how small a target the predator can detect.
- 12. Bats navigate using reflected sound waves. This process, known as echolocation, allows these animals to "see" in the dark. To uncover objects, bats must first emit a series of sound pulses. These pulses travel outward and strike objects. The pulses are then reflected off the objects and return back to the bats. Detected by their large ears, the sounds are quickly analyzed by the brain's echolocation center. This analysis is so precise that the bat can locate moving fish through a critical analysis of the ripples produced at the water's surface. Flying Blind Mammals can hear higher frequencies than other creatures can because of the characteristic arrangement of tiny bones in their ears as well as the structure of their cochlea, or inner ear. Even among mammals, however, hearing ability varies widely. Among people, most young adults can detect sounds with frequencies between 20 hertz, or cycles per second, and 20 kilohertz (kHz). But even that highest tone is low by echolocation standards: The typical bat's sonar chirps exploit frequencies as high as 120 kHz, and bottlenose dolphins' calls include frequencies that range up to 150 kHz or so. Birds, however, can't produce or hear the high-pitched, short-wavelength sounds needed to track insect-size targets. The few birds that can echolocate use lower frequencies, and they do so only to navigate in the dark, says J. Jordan Price, a biologist at St. Mary's College of Maryland in St. Mary's City. Even that limited capability provides a benefit, however, because it enables members of those species to nest in caves and other places that aren't readily accessible to predators. With one exception, all birds known to echolocate are swiftlets. Birds in this group catch insects on the fly just as a bat does, but they do so in the daytime and track their prey by sight, says Price. Scientists have typically relied on characteristics beyond size, shape, and color to distinguish the members of one swiftlet species from those of another, simply because the birds have so few distinguishing features, he notes. Until recently, all swiftlets known to echolocate fell within the genus Aerodramus. Then, Price and his colleagues found a swiftlet in another genus—the pygmy swiftlet, Collocalia troglodytes—sitting on its nest, in complete darkness, about 30 m inside a cave on an island in the Philippines.
- 13. Evolution of the Eye
- 14. Evolution of the Eye: When evolution skeptics want to attack Darwin's theory, they often point to the human eye. How could something so complex, they argue, have developed through random mutations and natural selection, even over millions of years? If evolution occurs through gradations, the critics say, how could it have created the separate parts of the eye -- the lens, the retina, the pupil, and so forth -- since none of these structures by themselves would make vision possible? In other words, what good is five percent of an eye? Darwin acknowledged from the start that the eye would be a difficult case for his new theory to explain. Difficult, but not impossible. Scientists have come up with scenarios through which the first eye-like structure, a light-sensitive pigmented spot on the skin, could have gone through changes and complexities to form the human eye, with its many parts and astounding abilities. Through natural selection, different types of eyes have emerged in evolutionary history -- and the human eye isn't even the best one, from some standpoints. Because blood vessels run across the surface of the retina instead of beneath it, it's easy for the vessels to proliferate or leak and impair vision. So, the evolution theorists say, the anti-evolution argument that life was created by an "intelligent designer" doesn't hold water: If God or some other omnipotent force was responsible for the human eye, it was something of a botched design. Bilogists use the range of less complex light sensitive structures that exist in living species today to hypothesize the various evolutionary stages eyes may have gone through.
- 15. Here's how some scientists think some eyes may have evolved: The simple light-sensitive spot on the skin of some ancestral creature gave it some tiny survival advantage, perhaps allowing it to evade a predator. Random changes then created a depression in the light-sensitive patch, a deepening pit that made "vision" a little sharper. At the same time, the pit's opening gradually narrowed, so light entered through a small aperture, like a pinhole camera. Every change had to confer a survival advantage, no matter how slight. Eventually, the light-sensitive spot evolved into a retina, the layer of cells and pigment at the back of the human eye. Over time a lens formed at the front of the eye. It could have arisen as a double-layered transparent tissue containing increasing amounts of liquid that gave it the convex curvature of the human eye. In fact, eyes corresponding to every stage in this sequence have been found in existing living species. The existence of this range of less complex light-sensitive structures supports scientists' hypotheses about how complex eyes like ours could evolve. The first animals with anything resembling an eye lived about 550 million years ago. And, according to one scientist's calculations, only 364,000 years would have been needed for a camera-like eye to evolve from a light-sensitive patch.