

Recommandé
Contenu connexe
Tendances
Tendances (20)
Similaire à Motor system cortical control
Similaire à Motor system cortical control (20)
Plus de PS Deb
Plus de PS Deb (20)
Dernier
Dernier (20)
Motor system cortical control
- 2. Organization of Motor Nervous System
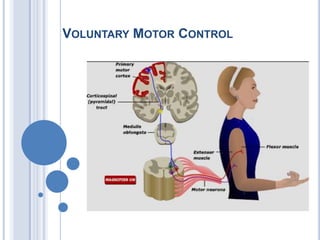
- 3. Control of voluntary movement Execution Planning Basal Ganglia Association cortex Premotor + Motor cortex Movement Idea Lateral cerebellum Intermediate Cerebellum
- 4. Organization of Motor system
- 5. Search for seat of Mind Ancient
- 6. Vitalism
- 7. The Renaissance of Neuroscience
- 8. Birth of NeuroAnatomy Leonardo Da Vinci 1452-1519 1664 - Thomas Willis Andreas Vesalius (1514-1564 )
- 9. Birth of NeuroPhysiology 1791 - Luigi Galvani 1809 - Luigi Rolando
- 10. Localizing Brain function Recent
- 11. Phrenology Franz Joseph Gall 1806 The theory of Gall and Spurzheim is ... an instance of a theory which, while essentially wrong, was just enough right to further scientific thought…Edwin Boring
- 12. 1824 - Marie-Jean-Pierre Flourens
- 14. 1825 - Jean-BaptisteBouillaud Presents cases of loss of speech after frontal lesions from clinicopathological correlation. We cannot test speech in animal Frontal lobe has other higher function which differentiate from animal. By symptom we can localize the cerebral lesion. Cerebellum controlled equilibrium and station Frontal lobe and higher mental functions
- 15. Hughling Jackson 1858 Three level of organization Spinal/brain stem Frontal Prefrontal Sensory integration and role of Basal ganglia was missed
- 16. Pierre Gratiolet <>Ernest Auburtin Brain Size and Volume brain acted as a whole and that localized functional centers did not exist
- 17. Lobar Localization Paul Broca1861
- 18. 1870, Hitzig and Fritsch Electrically stimulated various parts of a dog's motor cortex. They observed that depending on what part of the cortex they stimulated, a different part of the body contracted. Then they found that if they destroyed this same small area of the cortex, the corresponding part of the body became paralyzed. This is how it was discovered that every part of the body has a particular region of the primary motor cortex that controls its movement.
- 19. 1870 - David Ferrier
- 20. Discovery of Neuron RamonyCajal and Camillo Golgi 1906 Noble
- 21. 1906 - Sir Charles Scott Sherrington 1906- The Integrative Action of the Nervous system that describes the synapse and motor cortex Spinal reflex 1932 Nobel Prize
- 22. Leyton SSF & Sherrington CS (1917).Observations on the excitable cortex of the chimpanzee, orang-utan and gorilla. Q J Exp Physiol 11, 135–222. Figure 1. Motor maps of the gorilla cortex A, scale drawing of the left hemisphere of one of Leyton & Sherrington’s experiments on a gorilla (gorilla 1). The numbers and letters encode a wide range of different primary movements evoked by faradic stimulation. Eye movements (372–388) were generally evoked from an area further rostral from the motor cortex. Owing to lack of space, many motor effects were not plotted. B, simplified ‘map’ showing ‘responses grouped diagrammatically’,
- 24. Functional Organization of the Primary Motor Cortex
- 25. More intricate Motor Maps Intracortical micro-stimulation of layer V confirms the spatial motor map of Penfield Stimulation of small regions of the map activated single muscles, suggesting that vertical columns of cells in the motor cortex were responsible for controlling the actions of particular muscles. Microstimulation with simultaneous EMG recording shows that organized movements represented in motor map Individual pyramidal cells terminates on group of muscles in anterior horn cells in mosaic fashion
- 26. Lateral and Medial System The initiation of skilled voluntary movement in primates Loss of precise movement, retained power movement by lateral corticospinal destruction Anterior corticospinal system destruction produce axial muscle deficits that cause difficulty with balance, walking, and climbing.
- 27. Cortical control of movement 1947 Chang, Ruth could stimulate individual muscle by monkey motor cortex stimulation 1954 Bernhard and Bohm : single shock stimulation produces monosynaptic response in forelimb Landgren, Phillips and Porter (1962). stimulated the surface of the motor cortex while recording intracellularly from motoneurons and demonstrated the existante of a cluster of neurons which projected monosynaptically to motor neurons 1953, Malis, Pribram and Kruger showed that the motor cortex received afferent inputs from the periphery in the absence of the sensory cortex
- 28. Cellular organization of M1
- 29. The Organization of Cortical Efferent System
- 37. Experimental apparatus developed to record the activity of single neurons in awake primates trained to perform specific movements : Ed Evarts 1960
- 38. Direct corticospinal control of motor neurons is necessary for fine control of the digits
- 39. Motor Cortical Cell Firing with Force Generated
- 40. Corticomotoneuronal (CM) cell is active depends on the motor task
- 41. Activity in Individual Neurons of the Primary Motor Cortex Is Related to Muscle Force and Direction of Movement
- 42. Spike Triggered Averaging 1970
- 43. Different areas of cortex are activated during simple, complex, and imagined sequences of finger movements (Xenon PET)
- 44. Cell activity in the motor cortex depends on whether a sequence of movements is guided by visual cues or by prior training
- 45. A set-related neuron in the dorsal premotor area becomes active while the monkey prepares to make a movement to the left
- 46. The visuomotor transformations required for reaching and grasping involve two different pathways
- 47. Individual neurons in the ventral premotor area fire during specific hand actions only
- 48. Mirror Neurons A. Activity in the neuron as the monkey observes another monkey make a precision group.B. Activity in the same neuron as the monkey observes the human experimenter make the precision grip.C. Activity in the same neuron as the monkey itself performs a precision grip. (From Rizzolottiet al 1996.)
- 51. The Somatotopic Organization of the Motor Cortex Is Plastic
- 52. As a movement becomes more practiced, it is represented more extensively in primary motor cortex
- 53. Summary Primary Motor Cortex: Codes force and direction of movement Spinal motor neuron are directly under control for precise movement. Dorsal Premotor Cortex Movement related neuron encodes sensorimotor transformation for visual and sensory cue Fire before movement Ventral Premotor Cortex Encodes learned motor act fire before movement All cortical neurons are adaptable and plastic
- 54. Motor Cortex Afferent Adjacent cortex the somatosensory areas of the parietal cortex, theadjacent areas of the frontal cortex anterior to the motor cortex, and the visual and auditory cortices. Opposite cerebral hemisphere. Somatosensory fibers directly from the ventrobasal complex of the thalamus. Tracts from the ventrolateral and ventroanterior nuclei of the thalamus, which in turn receive signals from the cerebellum and basal ganglia Fibers from the intralaminar nuclei of the thalamus (RAS).
- 55. Convergence of Motor Control on the Anterior Motor Neuron
Notes de l'éditeur
- Commands for voluntary movement originate in cortical association areas. The movements are planned in the cortex as well as in the basal ganglia and the lateral portions of the cerebellar hemispheres, as indicated by increased electrical activity before the movement. The basal ganglia and cerebellum both funnel information to the premotor and motor cortex by way of the thalamus. Motor commands from the motor cortex are relayed in large part via the corticospinal tracts to the spinal cord and the corresponding corticobulbar tracts to motor neurons in the brain stem. However, collaterals from these pathways and a few direct connections from the motor cortex end on brain stem nuclei, which also project to motor neurons in the brain stem and spinal cord. These pathways can also mediate voluntary movement. Movement sets up alterations in sensory input from the special senses and from muscles, tendons, joints, and the skin. This feedback information, which adjusts and smoothes movement, is relayed directly to the motor cortex and to the spinocerebellum. The spinocerebellum projects in turn to the brain stem. The main brain stem pathways that are concerned with posture and coordination are the rubrospinal, reticulospinal, tectospinal, and vestibulospinal tracts and corresponding projections to motor neurons in the brain stem. Ganong 21st
- Spinal cord reflex mechanisms involve lower motor neurons and are subject to supraspinal control. The brain regions that have direct control over the spinal cord include the brainstem and cerebral cortex. Brainstem pathways that project to the spinal cord include the reticulospinal, vestibulospinal, rubrospinal, and tectospinal tracts. The cerebral cortex gives rise to both corticospinal and corticobulbar fibers. Corticospinal fibers are essential for voluntary control over fine movements, mainly of the distal extremities. Corticobulbar fibers contribute to the control of spinal cord indirectly, by acting on neurons of the brainstem that project to the spinal cord. Other corticobulbar fibers innervate lower motor neurons of the brainstem (cranial nerves) and provide the substrate and mechanism for voluntary movements of the head region. Two other regions, the basal ganglia and cerebellum, play important roles in motor functions. The basal ganglia affect motor systems by acting on neurons in the precentral and premotor regions that comprise the larger part of the corticospinal tract. The cerebellum affects motor function by acting on neurons in both the brainstem and the cerebral cortex that directly control motor functions of the spinal cord.
- In the late 1820s Gall's ideas were subjected to experimental analysis by the French physiologist Pierre Flourens. By systematically removing Gall's functional centers from the brains of experimental animals, Flourens attempted to isolate the contributions of each “cerebral organ” to behavior. From these experiments he concluded that specific brain regions were not responsible for specific behaviors, but that all brain regions, especially the cerebral hemispheres of the forebrain, participated in every mental operation. Any part of the cerebral hemisphere, he proposed, was able to perform all the functions of the hemisphere. Injury to a specific area of the cerebral hemisphere would therefore affect all higher functions equally.In 1823 Flourens wrote: “All perceptions, all volitions occupy the same seat in these cerebral) organs; the faculty of perceiving, of conceiving, of willing merely constitutes therefore a faculty which is essentially one.” The rapid acceptance of this belief (later called the aggregate-field view of the brain) was based only partly on Flourens's experimental work. It also represented a cultural reaction against the reductionist view that the human mind has a biological basis, the notion that there was no soul, that all mental processes could be reduced to actions within different regions in the brain!Eur Neurol. 2009;61(5):311-4. Epub 2009 Mar 17.Marie-Jean-Pierre Flourens (1794-1867) and cortical localization.Pearce JM.Department of Neurology, Hull Royal Infirmary, Hull, UK.AbstractThe child prodigy Marie-Jean-Pierre Flourens received his medical degree at Montpellier when aged 19. As a young promising physician Flourens was asked to investigate Gall's controversial views on cerebral localization. To test Gall's assertions, Flourens developed ablation as a procedure to explore the workings of the brain. By removing anatomically defined areas of the brain of an animal and watching its behaviour, he thought he might localize certain functions. Flourens did not favour the idea of cerebral localization and concluded that the brain functioned as a whole and thus arose the concept of 'cerebral equipotentiality'. This culminated in his 1824 Recherchesexpérimentalessur les propriétés et les fonctions du systèmenerveux. His techniques were, however, crude and imperfect, and his experiments were mainly on birds. Much criticism and debate ensued. A gifted man, Flourens also advanced the physiology of the vestibular apparatus and described the anaesthetic properties of ether.IntroductionBorn on 15 April 1794, in Maureilhan, near Béziers, France, Marie-Jean-Pierre Flourens (fig. 1) was something of a child prodigy. He enrolled at the famed Faculté de Médecine at Montpellier when he was only 15 years old and received his medical degree aged 19. Shortly thereafter, while Franz Gall (1758–1828) was at the height of his career in Paris, Flourens himself moved to the capital and under Georges Cuvier started to investigate brain functions and localization [1]. Gall’s work was questioned in some circles, and the Paris Academy of Sciences, acting on order of the Emperor Bonaparte, asked Flourens to investigate the matter. On the basis of his 1824 Recherchesexpérimentalessur les propriétés et les fonctions du systèmenerveux [2], he was elected to membership and eventually on Cuvier’s dying request was appointed Perpetual Secretary of the Académie des Sciences, to become one of France’s most revered scientists (fig. 2).Before Flourens began his animal experiments, Gall had developed relatively new ideas of brain function [3], which proved important:‘that the cerebral cortex represented the highest level of the brain and that the development of this area characterised mammals and man.’He tried to analyze and localize cerebral functions physiologically. His work moved ‘away from the concept of the insubstantial Cartesian soul to the concept of more material nerve function’, and in doing so ‘was wrong only in detail and in respect of the enthusiasm of its supporters’ [4]. His later ideas of phrenology [5] brought much of his work into disrepute, and physicians neglected his earlier significant advances. Nevertheless, as Edwin Boring wrote: ‘The theory of Gall and Spurzheim is ... an instance of a theory which, while essentially wrong, was just enough right to further scientific thought’ [6]. To test Gall’s assertions, Flourens developed ablation as a procedure to explore the workings of the brain. By removing anatomically defined areas of the brain of an animal and watching its behaviour, he thought he might localize certain functions. For example, by removal of the cerebellum, the animal could no longer coordinate movements. In 1823, he stated that the cerebellum regulates motor activity and a year later provided details of his ablation experiments on birds and lower vertebrates to study behaviour. By making discrete lesions in the medulla, near the posterior ventricle, he reported (cited in Changeux, p. 16, [7]):‘he disturbed certain vital functions, such as respiration.’Flourens did not favour the idea of cerebral localization and concluded that the brain functioned as a whole when he wrote:‘a large section of the cerebral lobes can be removed without loss of function. As more is removed, all functions weaken and gradually disappear. Thus the cerebral lobes operate in unison for the full exercise of their functions ... The cerebral cortex functioned as an indivisible whole ... [housing] an ‘‘essentially single faculty’’ of perception, judgement and will ... the last refuge of the soul’ (Flourens, cited by Changeux, p. 17, [7]).However, Flourens’s experimental technique was not accurate, and his behavioural studies were both crude and rudimentary for the adequate investigation of the faculties recently catalogued by Gall and Johann Spurzheim (1776–1832). Flourens related loss of function to the extent of damage. However, he adopted a holistic concept, with an understanding that the brain was composed of areas of specific functions, but specific areas interacted to work as a whole:‘All sensations, all perceptions, and all volition occupy concurrently the same seat in these organs. The faculty of sensation, perception, and volition is then essentially one faculty’ [2].Thus arose the concept of ‘cerebral equipotentiality’.While previous researchers had lesioned the brain through a trephine that made it impossible to localize damage accurately or to track haemorrhage, Flourens completely uncovered and isolated that portion of the brain to be removed. He tried to minimize operative trauma and post-operative bleeding and infection which might confound analysis. His ablation experiments led him to conclude the presence of a motor centre in the medulla oblongata and coordination in the cerebellum. Although his treatment of sensation was still rather confused, by the time the second edition of the Recherchesexpérimentales (1842) appeared, Flourens had clearly distinguished sensation and perception, treating perception as the appreciation of the meaning of a sensation. He thought that several sub-cortical structures were responsible for sensory function. However, his slicing of the hemispheres produced diffuse damage with resulting impairments of varied degree involving perception, intellect and will. The resulting damage varied with the extent of the lesion. After small lesions function might be restored, but total ablation caused permanent loss of function. Flourens concluded that while sensory-motor functions were localized to sub-cortical structures, higher mental processes were represented diffusely in the brain, all working as one.Gall and Flourens were vehemently opposed to each other’s work and ideas. Gall noted that Flourens’s procedure ‘mutilates all the organs at once, weakens them all, extirpates them all at the same time’. In fact, ablation by successive slices was a poor technique for demonstrating cortical localization and he was rightly reluctant to deduce general conclusions from lower organisms to primates and man. Flourens [8] attacked Gall’s efforts at localization and in contrast founded his theory of cerebral homogeneity that anticipated Lashley’s [9] much later concept of mass action and cortical equipotentiality. However, Flourens did concede that Gall had adduced detailed evidence of correlation between function and structure within the brain, and had established that the brain was the organ of mind. This was to be Gall’s most important contribution.Further DevelopmentsClinical localization was propounded by Jean-BaptisteBouillaud (1796–1881) [10] and Auburtin [11]. The psychological studies both of Alexander Bain and Herbert Spencer influenced the many advances of Hughlings Jackson.Precise localization remained, however, in dispute. On 4 April 1861, the Sociétéd’Anthropologie held a meeting. Paul Broca [12, 13] provided his now famous communication, which related partial destruction of the left frontal lobe to aphasia (aphemia) [14]. Jackson developed the concept that motor and sensory functions must be represented in an organized, hierarchical fashion. Fritsch and Hitzig, in 1870, showed in the dog that circumscribed cortical areas controlled movements of the contralateral limbs and that ablation caused weakness. Their findings confirmed the clinical localization of motor function. Testing Jackson’s notions, David Ferrier [15] used both ablation and electrical stimulation to produce topographical localization of function in several species. Published in his acclaimed The Functions of the Brain, Ferrier [16] had thereby mapped sensory and motor areas across several species, thus expanding both understanding and localization of movement and sensation [17].A celebrated debate in 1875 at the Société de Biologie in Paris between Charcot and Brown-Séquard illustrated the controversies of the day [18]. Charcot held an anatomical approach to localization of function. He relied on human pathological material primarily from isolated cerebral hematomas to establish the role of the cortex and subcortical white matter fibre tracts to motor and sensory function. By contrast, Brown-Séquard held a physiologist’s opinions, which disagreed fundamentally, arguing that since the brain consisted of complex networks, isolated lesions had no direct bearing on the localization of cerebral function. Though Brown-Séquard’s approach was to be vital in the evolving 20th-century concepts of brain function, it is generally agreed that in 1875, Charcot won the debate.In 1909, KorbinianBrodmann [19], stimulated by Oskar Vogt, described the different cytoarchitectonic structure of the gyri and showed that the human cortex is organized anatomically in the same way in man and other mammals. Brodmann’s discrete cortical areas [20] were, however, scorned with arguable justification by Henry Head. Other ContributionsFlourens [21] made other discoveries. In 1825 he observed that when a pigeon’s horizontal semi-circular canal was destroyed, the bird continued to turn in a circle, thus demonstrating the function of the vestibular labyrinth. He also showed that since destroying the medulla oblongata caused death of his animals, the medulla was responsible for circulation and breathing. In 1847, he described the anaesthetic properties of ether [22].Amongst Flourens’s pupils were EdméFélix Alfred Vulpian (1826–1887) and Gabriel Gustav Valentin (1810–1883). In 1840 he was elected to the French Academy, in preference to Victor Hugo. In 1845 he was created a commander of the légiond’honneur, and in the following year a peer of France. With the revolution in 1848 he withdrew from political life and in 1855 accepted the Professorship of Natural History at the Collège de France. He died at Montgeron, near Paris, on 6 December 1867.
- Bouillaud was born in 1796, studied medicine under Dupuytren and Magendie, being inspired by the latter with a profound interest in physiology. He received his doctorate in i823, and at the age of twentynine, published a brilliantmonograph, entitled "Traite clinique et physiologique de l'encephalite." In this volume he presented a number of clinical and pathological studies on loss of articulate speech associated with lesions of the anterior lobes of the brain, giving his reasons for localization of this function of the brain. In May of the same year, a further contribution was made, demonstrating that the loss of articulate speech corresponded to a lesion of the anterior lobes of the brain. These astute clinical studies won for him election to the Academy of Medicine in 1826, at the age of thirty. Four years later, he became Professor of Medicine, succeeding Recamier, joining the staff of the Charit6, in 1848, becoming Dean of the Faculty of Medicine, and in 1862 President of the Academy of Medicine.Bouillaud described cases from the literature and from his own practice showing that those who had lost the faculty of speech had lesions in the anterior lobes at postmortem. He made it clear that speech could be lost as an isolated functional defect and that loss of speech was not due to inability to move the tongue or lips, but to a lesion in the anterior lobes of the brain. Bouillaud's conclusions that the center for the faculty of speech resides in these lobes were based on clinical and pathological grounds.In support of his view, Bouillaud offered evidence showing that involvement of the anterior lobes alone was associated with loss of the faculty of speech. These he termed "positive arguments." Others were presented in which the disease affected other parts of the brain and in which speech was not affected. These he called "negative arguments." He reasoned that: "If it is true that the anterior lobes preside over speech, how can this be established since animals do not possess speech and therefore physiological experiments, on animals which have so widely contributed to science could furnish no positive evidence. In fact, man is the only animal who enjoys the noble privilege of speech; yet, man is sacred and inviolable for experimental physiology. Yet, disease, nature's own experiment on man, no less ingenious than the most skillful physiologist, can furnish the elements essential for the solution of our problem.How often nature, by means of illness, destroys the functions of the anterior lobes of the brain, the kind of a lesion which for our end is equivalent to ablation itself." "If on the one hand physiology is the true light of pathology, it is equally certain that pathology is for physiology an inexhaustible source of information. It leads us in a most direct manner to an understanding of the function of organs.“Bouillaud held that "the frontal lobes, in effect, preside over the great act of speech; but it is not by speech alone and the operations which accompany it, such as writing, arithmetic, etc., that man exercises supreme control over his environment, and extends indefinitely the range of his intelligence and the sphere of his perfectability. It is, therefore, probable that the anterior part of the brain is not exclusively devoted to the important functions of articulate language, but that it is also the seat of the highest functions of judgment. Let us -then cease to be astonished if the frontal area of man proves an extent proportionately so superior to the frontal region of all other animals; it could not be otherwise since in man, in that part of the brain corresponding to the frontal lobes, are situated the centres whose sublime gift he alone has received from nature.“Thus, not only does he place speech in the frontal lobes, but also the higher intellectual faculties. These revolutionary views concerning the functions of the anterior lobes, he repeatedly presented during the ensuing years, before the Academy of Medicine and the Academy of Science, based on both clinical and pathological material. Yet, in spite of the evidence offered, his views met with little acceptance.By his critical analyses of the clinical problems before him Bouillaud felt that the time had come when medicine could propose the following problem: "Given the symptoms, determine the seat of the disease," and conversely: "We shall see in fact, that symptoms vary according to the location of the cerebral affection; that muscular paralysis, for example, varies according to the location the lesion occupies, whether the anterior, middle or posterior lobes of the brain, and we shall by this method be able to determine some of the functions devolved upon various parts of which the cerebrum is composed. From this last point of view one of the most interesting results, to which clinical observations have led us, is that the anterior part of the brain is truly the centre of articulated language.“Bouillaud reasoned further that a focal and limited paralysis must indicate a focal and limited lesion of the brain, stating that: "The plurality of cerebral centres reserved for movement is in effect proven by the existence of isolated partial paralysis corresponding to a local alteration of the brain; since it is evident that if this organ were not made up of several motor or conductor centres for muscular movement, it would be impossible to conceive how a lesion of one of these areas carried with it a paralysis of a given part of the body without affecting movement of any of the other parts.“Bouillaud's contributions were not limited to the function of the cerebrum. In 1827 he published the results of his experimental andclinical studies indicating that the cerebellum controlled equilibrium and station, refuting Gall's view that the cerebellum was an organ of the instinct of propagation, thus recognizing important functions of the cerebellum.Thus, the basic principles overthrowing the concept that the brain acted as a whole were solidly laid by Bouillaud in 1825, and on numerous subsequent occasions, notably in i839 and in 1848. Unfortunately, his monumental contributions in the field of neuro-anatomy and clinical neurology have not even yet been generally recognized; whereas, his work on the heart and the relationship of rheumatic fever to endocarditis have been generally appreciated.*The opposition with which Bouillaud's views on cerebral localization were met seems hard to understand since they were adequatelysupported by clinical and pathological evidence; yet in spite of these clinical and pathological contributions indicating the presence of various centers in the brain for specific and localized functions the problem remained unsettled. We find a great debate concerning cerebral localization and the existence of cerebral centers being carried on in the Anthropological Society of Paris during i86i, some thirty-six years after Bouillaud's original contributions.
- As originally conceived by Hughlings Jackson in 1858, purely on the basis of clinical observations, the motor system is organized hierarchically in three levels, each higher level controlling the one below. It was Jackson’s concept that the spinal and brainstem neurons represent the lowest, simplest, and most closely organized motor centers; that the motor neurons of the posterior frontal region represent a more complex and less closely organized second motor center; and that the prefrontal parts of the cerebrumare the third and highest motor center. This scheme is still regarded as being essentially correct, though Jackson failed to recognize the importance of the parietal lobe and basal ganglia in motor control.Since Jackson’s time, physiologists have repeatedly analyzed these three levels of motor organization and have found their relationships to be remarkably complex. Motor and sensory systems, although separated for practical clinical purposes, are not independent entities but are closely integrated. Without sensory feedback, motor control is ineffective.And at the higher cortical levels of motor control, motivation, planning, and other frontal lobe activities that subserve volitional movement are always preceded and modulated by activity in the parietal sensory cortex.Motor activities include not only those that alter the position of a limb or other part of the body (isotonic contraction) but also those that stabilize posture (isometric contraction).Movements that are performed slowly are called ramp movements. Very rapid movements are called ballistic (they are too fast for sensory control). Another way of classifying movements, stressed by Hughlings Jackson, is in terms of their automaticity: reflex movements are the most automatic, willed movements the least. Physiologic studies, cast in their simplest terms, indicate that the following parts of the nervous system are engaged primarily in the control of movement and, in the course of disease, yield a number of characteristic derangements.
- During February, March, and April, 1861, a notable series of meetings of the Anthropological Society took place in Paris, in which * At Bouillaud's request, no oration was held at his funeral; however, four years later his compatriots, recognizing his many contributions and the dominant role which he had played in medicine, erected a statue of him au Place Angouleme.Pierre Gratiolet, Ernest Auburtin and Paul Broca participated, which culminated finally in the recognition and establishment of the principle of cerebral localization.This was a great debate, a continuing debate, carried on primarily by Pierre Gratiolet, the fame of whose contribution in anatomy carried such tremendous weight that it made any opposing opinion difficult to sustain. Nevertheless, undaunted, Ernest Auburtin, thirty-six years old, Chief of Clinic at Charite, devoted pupil and son-in-law of Bouillaud, repeatedly rose in the various meetings to disagree with Gratiolet's concept that the brain acted as a whole and that localized functional centers did not exist. Auburtin insisted that were one single localized cerebral function established, then the principle of cerebral localization would be demonstrated. Were the principle of cerebral centers proven for one function, the centers controlling other functions could then be unravelled and the brain elevated to its proper position, no longer conceived of as acting as a single unit, as it had for centuries been thought to do.Auburtin held that if it can be demonstrated that lesions which abolish speech constantly occupy a definite cerebral convolution onecan hardly fail to admit that this convolution is the seat of articulated language. Proof of the existence of such a center of localized function would become the key to unlock additional centers.Gratiolet's paper at these meetings had to do with the size, volume, etc., of the brain as a whole from an anthropological viewpoint. He presented a Tolenac skull and indicated the importance of the volume of the brain, holding that size and weight were the determining factors in intelligence and firmly expressing his belief that the brain acted as a whole, that all parts of the brain participated simultaneously and equally in intelligence.Broca also participated in these meetings. His paper was entitled "On the Volume and Form of the Brain According to Individuals andAccording to Race," thus carrying on in general the concept of Gratiolet that the size and shape of the brain determined its function as a whole.However, Broca felt that separation of the brain into anatomical divisions also meant separation of function, holding that we already had enough evidence to say that all cerebral convolutions do not have the same attributes, maintaining that in the brain, as pointed out by Gall and Spurzheim, there are distinct functions associated with anatomical division. Broca, however, while leaning toward the concept of cerebral localization did not come to the point of identifying any particular function with any particular part of the brain.Auburtin, opposing the views of Gratiolet, insisted that the opinion first expressed by Gall, and subsequently elaborated by Bouillaud,namely that the faculty of speech was to be found in the anterior lobe had been adequately demonstrated clinically. Auburtin agreed that the brain was a complex organ with multiple and diverse functions but that greater advances would be made, and more precise information obtained, were the brain considered in its various parts, rather than as a whole.Auburtin argued, as did Bouillaud before him, that the highest cerebral functions reside in the anterior lobes and are in proportion to the development of these lobes. To this Gratiolet replied that research thus far had failed to find any localization of faculties in the brain. However, Auburtin called attention to the fact that various brain lesions produced quite different signs, depending upon their localization. The lesions of the anterior lobe produce neither loss of sensation nor of motion, yet abolish the faculty of speech. Gratiolet held that the difference depended upon whether the lesion occurred slowly or rapidly. To this, Auburtin answered that while different pathological lesions may produce different results, it is none the less important that similar lesions in simikir areas produce similar results.Auburtin insisted on the importance of examining a particular function of the anterior lobes in order that the problem might be brought into sharper focus. He then called attention to a number of cases cited in the literature, notably those by Lallerman, Rosten, and particularly those in Bouillaud's "Traite de l'encephalite," in which the clinical findings of loss of the faculty of articulate speech had permitted them to predicate and subsequently demonstrate at autopsy a lesion in the frontal lobes. Auburtin then reported a most striking clinical observation and clinical experiment which has been curiously neglected as one of the first instances of determining physiological data upon the human brain with the patient conscious and able to cooperate in the experiment. A patient who had tried to commit suicide by shooting himself through the frontal region, entered l'Hopital St. Louis. A part of the frontal bone was removed, exposing the frontal lobe which was not lacerated. The patient's intelligence was intact, as well as his speech. He survived sufficiently long to be the subject of a very interesting experiment. When a spatula was applied to the anterior lobe, gently without causing general pressure, speech would instantly be abolished, only to return as soon as the spatula was removed. This was done repeatedly, with repeated loss of the faculty of speech unaccompanied by any other paralysis. Its clinical implications and its significance were realized by Auburtin.Auburtin recognized that the anterior lobes were large and that, as yet, the precise area where the faculty of speech resides had not been determined. Instances could be cited in which a lesion existed in one anterior lobe and not the other, suggesting that perhaps one being intact, the intact lobe is able to contribute in part to the function of the other lobe.In order to bring the discussion which had taken place at length to a final conclusion, Auburtin on April 4, challenged those who opposed cerebral localization by saying that "On the service of M. Bouillaud, I have studied for a long time a patient named Bache who has lost his speech, who, nevertheless understands all that is said to him, replying by signs in an intelligent manner to all questions put to him. This patient has spent several years at Bicetre and is now at the Hospital for Incurables.I have seen him recently, his illness has progressed, a slight paralysis has supervened, but his intelligence is still intact and his speech still abolished. This man will die without doubt, in a short time. In view of the symptoms which he presents, I have made a diagnosis of softening of the anterior lobes. If at autopsy the anterior lobes are found intact, then I shall renounce the ideas which I have sustained, but I can only reason in accordance with the facts which today exist in science. Still, I have never seen a lesion limited to the middle or posterior lobe destroy the faculty of speech.“To this Gratiolet replied that he could cite cases in which the frontal lobes were involved without loss of speech and replied: "I do not hesitate to conclude that all attempts at localization, which up to now have been tried, lack any foundation. They are no doubt great efforts, titanic efforts! But when one attempts to grasp the truth at the height of these babbles, the edifice crumbles." It is noteworthy that throughout this debate in which Gratiolet did not hesitate to be emphatic in his views concerning the fallacy of cerebral localization and Auburtin equally vigorously attacking the concept that the brain acted as a whole, as a single unit, Broca though present at each of the sessions did not commit himself as to the fundamental concept concerning the relationship of the faculty of speech and the anterior lobes.
- Electrically stimulated various parts of a dog's motor cortex. They observed that depending on what part of the cortex they stimulated, a different part of the body contracted. Then they found that if they destroyed this same small area of the cortex, the corresponding part of the body became paralyzed. This is how it was discovered that every part of the body has a particular region of the primary motor cortex that controls its movement
- David FerrierIn 1870, two German scientists, Gustav Fritsch and Eduard Hitzig, reported their findings on the electrical excitability of the exposed cerebral cortex in dogs in which galvanic current was used. According to Horwitz,[5] David Ferrier was impressed by these scientists' work, and he refined their techniques. By using faradic current stimulation, Ferrier investigated even further the cortical functioning of many different animals, including primates. He constructed one of the first detailed cortical maps and confirmed many of the principles set forth by Dr. Jackson. Ferrier firmly established the location of the motor cortex, stating that it extended along the rolandic fissure medially to the interhemispheric area. Not all of his conclusions were correct, however. For example, he erroneously placed the center for vision in the superior temporal lobes, and similarly was unable adequately to determine the function of the occipital lobes from his studies. Ferrier summarized his results in the 1876 publication of The Functions of the Brain ,[4] and dedicated this work to Jackson, stating "To Dr. Hughlings Jackson, who from a clinical and pathological standpoint anticipated many of the more important results of recent experimental investigation into the functions of the cerebral hemispheres." As discussed in Horwitz,[5] 2 years later, Ferrier produced a second publication titled The Localisation of Cerebral Disease , and he dedicated it to a well-known French neurologist, Dr. Jean-Martin Charcot.
- Charles Scott Sherrington was born on November 27, 1857, at Islington, London. He was the son of James Norton Sherrington, of Caister, Great Yarmouth, who died when Sherrington was a young child. Sherrington's mother later married Dr. Caleb Rose of Ipswich, a good classical scholar and a noted archaeologist, whose interest in the English artists of the Norwich School no doubt gave Sherrington the interest in art that he retained throughout his life.In 1876 Sherrington began medical studies at St. Thomas's Hospital and in 1878 passed the primary examination of the Royal College of Surgeons, and a year later the primary examination for the Fellowship of that College. After a short stay at Edinburgh he went, in 1879, to Cambridge as a noncollegiate student studying physiology under Michael Foster, and in 1880 entered Gonville and Caius College there.In 1881 he attended a medical congress in London at which Sir Michael Foster discussed the work of Sir Charles Bell and others on the experimental study of the functions of nerves that was then being done in England and elsewhere in Europe. At this congress controversy arose about the effects of excisions of parts of the cortex of the brains of dogs and monkeys done by Ferrier and Goltz of Strasbourg. Subsequently, Sherrington worked on this problem in Cambridge with Langley, and with him published, in 1884, a paper on it. In this manner Sherrington was introduced to the neurological work to which he afterwards devoted his life.In 1883 Sherrington became Demonstrator of Anatomy at Cambridge under Professor Sir George Humphrey, and during the winter session of 1883-1884 at St. Thomas's Hospital he demonstrated histology.The years 1884 and 1885 were eventful ones for Sherrington, for during the winter of 1884-1885 he worked with Goltz at Strasbourg, in 1884 he obtained his M.R.C.S., and in 1885 a First Class in the Natural Sciences Tripos at Cambridge with distinction. During this year he published a paper of his own on the subject of Goltz's dogs. In 1885 he also took his M.B. degree at Cambridge and in 1886 his L.R.C.P.In 1885 Sherrington went, as a member of a Committee of the Association for Research in Medicine, to Spain to study an outbreak of cholera, and in 1886 he visited the Venice district also to investigate the same disease, the material then obtained being examined in Berlin under the supervision of Virchow, who later sent Sherrington to Robert Koch for a six weeks' course in technique. Sherrington stayed with Koch to do research in bacteriology for a year, and in 1887 he was appointed Lecturer in Systematic Physiology at St. Thomas's Hospital, London, and also was elected a Fellow of Gonville and Caius College, Cambridge. In 1891 he was appointed in succession to Sir Victor Horsley, Professor and Superintendent of the Brown Institute for Advanced Physiological and Pathological Research in London. In 1895 he became Professor of Physiology at the University of Liverpool.During his earlier years in Cambridge, Sherrington, influenced by W. H. Gaskell and by the Spanish neurologist, Ramón y Cajal, whom he had met during his visit to Spain, took up the study of the spinal cord. By 1891 his mind had turned to the problems of spinal reflexes, which were being much discussed at that time, and Sherrington published several papers on this subject and, during 1892-1894, others on the efferent nerve supply of muscles. Later, from 1893-1897, he studied the distribution of the segmented skin fields, and made the important discovery that about one-third of the nerve fibres in a nerve supplying a muscle are efferent, the remainder being motor.At Liverpool he returned to his earlier study of the problem of the innervation of antagonistic muscles and showed that reflex inhibition played an important part in this. In addition to this, however, he was studying the connection between the brain and the spinal cord by way of the pyramidal tract, and he was at this time visited by the American surgeon Harvey Cushing, then a young man, who stayed with him for eight months.In 1906 he published his well-known book: The Integrative Action of the Nervous System, being his Silliman Lectures held at Yale University the previous year, and in 1913 he was invited to become Waynfleet Professor of Physiology at Oxford, a post for which he had unsuccessfully applied in 1895, and here he remained until his retirement in 1936. Here he wrote, and published in 1919, his classic book entitled Mammalian Physiology: a Course of Practical Exercises, and here he regularly taught the students for whom this book was written.In physique Sherrington was a well-built, but not very tall man with a strong constitution which enabled him to carry out prolonged researches.During the First World War, as Chairman of the Industrial Fatigue Board, he worked for a time in a shell factory at Birmingham, and the daily shift of 13 hours, with a Sunday shift of 9 hours, did not, at the age of 57, tire him. From his early years he was short-sighted, but he often worked without spectacles.The predominant notes of his character as a man were his humility and friendliness and the generosity with which he gave to others his advice and valuable time. An interesting feature of him is that he published, in 1925, a book of verse entitled The Assaying of Brabantius and other Verse, which caused one reviewer to hope that «Miss Sherrington» would publish more verse. He was also sensitive to the music of prose, and this and the poet in him, but also the biologist and philosopher, were evident in his Rede Lecture at Cambridge in 1933 on The Brain and its Mechanism, in which he denied our scientific right to join mental with physiological experience.The philosopher in him ultimately found expression in his great book, Man on his Nature, which was the published title of the Gifford Lectures for 1937-1938, which Sherrington gave. As is well known, this book, published in 1940, centres round the life and views of the 16th century French physician Jean Fernel and round Sherrington's own views. In 1946 Sherrington published another volume entitled The Endeavour of Jean Fernel.Sherrington was elected a Fellow of the Royal Society of London in 1893, where he gave the Croonian Lecture in 1897, and was awarded the Royal Medal in 1905 and the Copley Medal in 1927. In 1922 the Knight Grand Cross of the Order of the British Empire and in 1924 the Order of Merit were conferred upon him. He held honorary doctorates of the Universities of Oxford, London, Sheffield, Birmingham, Manchester, Liverpool, Wales, Edinburgh, Glasgow, Paris, Strasbourg, Louvain, Uppsala, Lyons, Budapest, Athens, Brussels, Berne, Toronto, Montreal, and Harvard.As a boy and a young man Sherrington was a notable athlete both at Queen Elizabeth's School, Ipswich, where he went in 1871, and later at Gonville and Caius College, Cambridge, for which College he rowed and played rugby football; he was also a pioneer of winter sports at Grindelwald.In 1892 Sherrington married Ethel Mary, daughter of John Ely Wright, of Preston Manor, Suffolk. After some years of frail health, during which, however, he remained mentally very alert, he died suddenly of heart failure at Eastbourne in 1952.From Nobel Lectures, Physiology or Medicine 1922-1941, Elsevier Publishing Company, Amsterdam, 1965 This autobiography/biography was written at the time of the award and first published in the book series Les Prix Nobel. It was later edited and republished in Nobel Lectures. To cite this document, always state the source as shown above.
- Figure 1. Motor maps of the gorilla cortexA, scale drawing of the left hemisphere of one of Leyton & Sherrington’s experiments on a gorilla (gorilla 1). The numbers and letters encode a wide range of different primary movements evoked by faradic stimulation. Eye movements (372–388) were generally evoked from an area further rostral from the motor cortex. Owing to lack of space, many motor effects were not plotted. B, simplified ‘map’ showing ‘responses grouped diagrammatically’, as the authors put it.The paper by Leyton & Sherrington (1917) has had a profound and lasting impact for ninety or so years since its publication.It is notable in many different ways. Leyton & Sherrington (1917) provided the first detailed proof that there was indeed localization of function within the cerebral cortex. The durability of their report probably owes most to the fact that Leyton & Sherrington (1917)were the first to establish precisely the true extent of the motor area, and to provide the first detailed ‘motor map’ of the primate motor cortex. In addition, they showed that surgical extirpation of the cortical tissue that, when stimulated, gave rise to movement of a particular body part, resulted in a widespread weakness and loss of use of that same body part.There was, however, substantial recovery in the weeks that followed, recovery that was not lost on lesioning either the adjacenttissue in the same hemisphere or the equivalent cortical area of the opposite hemisphere. Finally, they were able to trace the course of the degenerating corticofugal and corticospinalfibres. They observed widespreaddegeneration in the cervical cord after a lesion of the hand and arm cortical area and noted that after such a lesion in the chimpanzee (p. 185), ‘the whole of the cross-area of ventral horn has scattered through it many degenerating fibres. . .’, which I think is the first report of the direct cortico-motoneuronal projection, a projection whose existence was confirmed physiologically by Bernhard & Bohm (1954), and one that appears to be unique to primates (Porter & Lemon, 1993).The mysterious chronology of the paperThis classic paper was very long in gestation. After the early accounts appeared in 1901 and in 1903, further details of their experiments were given in Sherrington’s Silliman Lectures (published as The Integrative Action of the Nervous System in 1906). However, the full paper was not submitted to the Quarterly Journal of Physiology until October 1916. There is a much-repeated story, probably first promulgated by John Fulton in his 1952 biography, that Sherrington delayed publication until after the death of Victor Horsley, with whom he had a very heated row. It was believed that Sherrington wanted to avoid a repeat of the disagreement in 1894 over primacy of research on pyramidal tract degeneration. Horsley, the famous neurosurgeon and co-inventor the Horsley–Clark stereotaxic method, died in July 1916 from fatal sunstroke in Mesopotamia (where he was stationed during the War).However, this account has been seriously questioned (Vilenskyet al. 2003) and there must have been other reasons for the delay; these might have included the expense and difficulty of obtaining the additional animals needed for the study, and the facts that Leyton left Liverpool not long after the 1903 paper was published, to take up a Chair in Leeds, and that the the work was undoubtedly interrupted by the outbreak of the GreatWar. Why was the study carried out?The decision to investigate the excitable cortex of anthropoid apes probably arose because the findings of motor responses evoked by cortical stimulation in animals such as dogs, rabbits and monkeys were challenged as of being of little relevance to humans (where have we heard that before?).Sherrington considered that experiments in ‘higher apes’, being closer to humans than any other species, might provide the answer. The initial experiment (in a chimpanzee) having provided a positive result, they decided to complete a full series of studies, and this paper summarizes agreat many different experiments in a total of 22 chimpanzees, three orang-utans three gorillas and other animals, all investigated under deep anaesthesia with chloroform and ether. One of them involved an experiment in which the same induction coil was used to stimulate, simultaneously, the cortex of a cat, amonkey and a chimpanzee, and this demonstrated unequivocally that the threshold for evoking motor response was very similar in all three species.Rereading this paper, one is struck by it having all the hallmarks of a really modern piece of neuroscience: a multidisciplinary mixture of state-of-the art electrophysiology, behavioural analysis and detailed neuroanatomy, all linked together to test a strong central hypothesis: ‘the motor cortex can be regarded as a synthetic organ for compounding and recompounding in varied ways movements of varied kinds. . . . ’ The authors considered that the movements they evoked were like words in a basic vocabulary of voluntary movement, words that could be combined in myriad ways to make sentences: purposeful movements involving co-ordinated activation of many muscles in a precise spatio-temporal sequence.There are a great many key points of lasting value in this paper, some of which are discussed below.What can be deduced about brain motor function from electrical stimulation?Sherrington was very clear that the nature of the motor responses was in part due to the type of stimulation used. Leyton & Sherrington (1917) employed ‘faradic’ stimulation (alternating current provided by an induction coil), were in favour of unipolar stimulation (using a large andremote indifferent electrode) and insisted on studying responses evoked by a brief period of stimulation at the lowest current intensities. In this way they reduced the possibility that the responses were due to physical spread of the induced current through the tissue. Stronger and longer lasting stimulation also raises the possibility that the responses are due to physiological spread of synaptic activity generated locally at the stimulation site but permeating through both short- and long-distance connections to recruit even quite remote cortical and subcortical structures.As Phillips & Porter (1977), in their elegant discussion of Leyton & Sherrington (1917) put it: ‘Thus faradic stimulation is disqualified, equally with “galvanic” as tool for evoking natural function but it can be used as ‘a tool for mapping the outputs that are available for selection by the intracortical activities that it cannot itself evoke.’ Previous investigators, including Ferrier and Beevor & Horsley, had used relatively large currents and thus been able to evoke motor responses from the primary sensory as well as from the primary motor cortex.Leyton & Sherrington (1917) refined the approach by carefully controlling current intensity, and demonstrated that the motor area did not extend posterior to the central sulcus. They were careful to point out that even a motor zone could influence sensation, a prescient comment given thatmany of the descending corticospinalfibres are involved in control of afferent input (see Lemon & Griffiths, 2005; Lemon, 2008). The quality of the movements evoked by electrical stimulationLeyton & Sherrington (1917), describing the ‘fractional quality’ of the motor responses evoked, write that ‘the individual movements, elicited by somewhat minutely localised stimulations, are, broadly speaking, fractional, in the sense that each, though co-ordinately executed, forms,so to say, but a unitary part of some more complex act, that would, to attain its purpose, involve combination of that unitary movement with others to make up a useful whole.’ They were also keen to emphasize the complexity of some of the responses, which often comprised a‘1st movement’ (e.g. thumb extension) with accompanying 2nd (e.g. index finger extension), 3rd (wrist extension) or even 4th movements. They also stressed the ‘functional instability of cortical motor points’; the responses were not fixed, but showed properties such as facilitation,reversal and deviation, especially when a given cortical point was retested after the adjacent area had been stimulated. Finally, they recognized that movements of the eyes were likely to be controlled differently from movements of the rest of the body, since the area giving rise to eye movements was located rostral to and distinct from the main motor cortex (see Fig. 1), in what we now call the frontal eye fields.How many different kinds of movements can be evoked and how are they ‘mapped’ across the cortical surface?Leyton & Sherrington (1917) were clearly struck by the rich variety of different movements that could be evoked from the motor area. They reported more than 400 different ‘1st movements’ in a table that takes up 7 pages (would the current editors of Experimental Physiology allowsuch indulgence?). Each movement was carefully inscribed on a scale drawing of the exposed cortex (Fig. 1A). They were careful to note that the map is not fixed in nature but that responses showed instability in time (see above), and that even the boundaries between different body ‘areas’(e.g. between the face and arm area) were labile. Movement of a given body part (e.g. the thumb) could be evoked from an extensive region of motor cortex, many square millimetres in area, and this representation overlapped significantly with that of other, adjacent body parts (e.g.thumb with index finger, wrist with elbow etc.).Similar findings were published 20 years later by Penfield & Boldrey (1937) for the human brain. Somehow the complexity of the experimental findings was digested into a greatly simplified ‘homunculus’ published after another long delay by Penfield & Rasmussen (1952). This cartoon suggested a strictly somatotopic representation within the primary motor area, M1, which bore little relation to the original experimentaldata. However, the homunculus provided an appealing notion of how voluntary motor control might be represented in the cortex, and has since been reproduced in every relevant textbook. It often appears alongside Sherrington’s (1906) map of the gorilla cortex, which bears some resemblance to figure 10 of Leyton & Sherrington (1917), a figure which is not referred to at all in the text (more editorial indulgence?). Thisfigure, in which the responses were grouped diagrammatically (sic), is reproduced here as Fig. 1B. Its simplicity conceals the wealth of data that each of their mapping experiments yielded (Fig. 1A). While the orderly mapping of different areas of the body is well substantiated, evidence for a strict somatotopy within each area (e.g. fixed non-overlapping ordering in the lateral to medial direction of thumb, then index, middle, ring and little finger movement) has been hard to find, and modern investigators (see Lemon, 1988; Schieber&Hibbard, 1993;Rathelot&Strick, 2006) have turned away from this concept to one much closer to the multiple and overlapping representation of movement described by Leyton & Sherrington (1917). This type of representation is well suited for the many and varied combination of ‘fractional’ movements into useful actions or, as they put it: ‘. . .that from movements of locally restricted parts, e.g. movements of a finger or of a limb joint (movements themselves discrete andindividually separable in the motor cortex), the up building of larger combinations varied in character and serviceable for purposes of different and varied kind, prehensile, defensive, locomotor, mimetic, masticatory, deglutitional, orientational etc. is one of the main offices performed by themotor cortex.’ We are now learning how individual motor cortex neurons combine and recombine their actions (Jackson et al. 2003).Why is the cortex involved in motor control?Many investigators have raised the issue of why the basic building blocks of all types of movements are represented at all in the neocortex, one of brain’s ‘higher’ centres; surely these could be managed by subcorticalcentres, including the spinal cord? Leytonand Sherrington recognized, ofcourse, that the spinal cord could generate many types of movements. However, ‘It would seem that in order to preserve the possibility of being interchangeably compounded in a variety of ways, successive or simultaneous, these movements must lie, as more or less discrete and separableelements, within the grasp of the organ whichhas the varied compounding of them.’ Beautifully put! They also recognized that the acquisition of new combinations would make an important contribution to the learning of motor skills: ‘The acquirement of skilled movements, though certainly a process involving far wider areas of the cortex than the excitable zone itself, may be presumed to find in the motor cortex an organ whose synthetic properties are part of the physiological basis which renders that acquirement possible.’ConclusionLeyton and Sherrington’s seminal paper provided a key set of observations that have guided and inspired research ever since. They were clearly impressed by the degree of recovery after large cortical lesions and recognized that some kind of compensatory changes must be at work.They also recognized the importance of finding the neural substrate of recovery. It was only much later that it became clear that plasticity within the cortex is itself an active, use-dependent process, and that this process can be harnessed to improve recovery of function. Ultimately, Leyton and Sherrington’s work opened up an exciting new chapter in the physiology of the cortex, which ultimately is having far-reaching consequences for our understanding of the brain and for the rehabilitation of patients with brain injury.ReferencesBernhard CG & Bohm E (1954). Cortical representation and functional significance of the corticomotoneuronal system. AMA Arch Neurol Psychiatry 72, 473–502. Granit R (1966). Charles Scott Sherrington: An Appraisal. Thomas Nelson and Sons, London. Gr¨unbaum ASF & Sherrington CS (1901).Observations on the physiology of the cerebral cortex of some of the higher apes (preliminary communication). Proc Roy Soc Lond69, 206–209. Gr¨unbaum ASF & Sherrington CS (1903). Observations on the physiology of the cerebral cortex of the anthropoid apes. Proc Roy Soc Lond72, 152–155. Jackson A, Gee VJ, Baker SN & Lemon RN (2003). Synchrony between neurons with similar muscle fields in monkey motor cortex. Neuron 38, 115–125.Lemon RN (1988). The output map of the primate motor cortex. Trends Neurosci11, 501–506.Lemon RN (2008). Descending pathways in motor control. Ann Rev Neurosci; DOI: 10.1146/annurev.neuro.31.060407.125547.Lemon RN & Griffiths J (2005). Comparing the function of the corticospinal system in different species: organizational differences for motor specialization? Muscle Nerve 32, 261–279.Leyton SSF & Sherrington CS (1917). Observations on the excitable cortex of the chimpanzee, orang-utan and gorilla. Q J Exp Physiol11, 135–222.PenfieldW&Boldrey E (1937). Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 60, 389–443.PenfieldW&Rasmussen T (1952). The Cerebral Cortex of Man. Macmillan, New York. Phillips CG & Porter R (1977). Corticospinal Neurones. Their Role in Movement. Academic Press, London. Porter R & Lemon RN(1993). Corticospinal Function and Voluntary Movement. Clarendon Press, Oxford.Rathelot J-A & Strick PL (2006). Muscle representation in the macaque motor cortex: an anatomical perspective. Proc NatlAcadSci USA 103, 8257–8262.Schieber MH & Hibbard LS (1993). How somatotopic is the motor cortex hand area? Science 261, 489–492.Sherrington CS (1906). The Integrative Action of the Nervous System. Cambridge University Press.Stewart MJ (1922). Albert Sidney FrankauLeyton (obituary). J PatholBacteriol25, 109–112.Vilensky JA, Stone JL & Gilman S (2003). Feud and fable: the Sherrington–Horsley polemic and the delayed publication. J HistNeurosci12, 368–375.
- The introduction in the 1960s of intracorticalmicrostimulation (a more refined method of cortical activation) allowed a more detailed understanding of motormaps. Microstimulation entails the delivery of electrical currents an order of magnitude smaller than those used by Sherrington and Penfield. By passing the current through the sharpened tip of a metal microelectrode inserted into the cortex, the upper motor neurons in layer V that project to lower motor neuroncircuitry can be stimulated focally. Although intracortical stimulation generally confirmed Penfield's spatial map in the motor cortex, it also showed that the finer organization of the map is rather different than most neuroscientists imagined. For example, when microstimulation was combined with recordings of muscle electrical activity, even the smallest currents capable of eliciting a response initiated the excitation of several muscles (and the simultaneous inhibition of others), suggesting that organized movements rather than individual muscles are represented in the map (see Box B). Furthermore, within major subdivisions of the map (e.g., arm, forearm, or finger regions), a particular movement could be elicited by stimulation of widely separated sites, indicating that nearby regions are linked by local circuits to organize specific movements. This interpretation has been supported by the observation that the regions responsible for initiating particular movements overlap substantially.What Do Motor Maps Represent?Electrical stimulation studies carried out by the neurosurgeon Wilder Penfield and his colleagues in human patients (and by Clinton Woolsey and his colleagues in experimental animals) clearly demonstrated a systematic map of the body's musculature in the primary motor cortex (see text). The fine structure of this map, however, has been a continuing source of controversy. Is the map in the motor cortex a “piano keyboard” for the control of individual muscles, or is it a map of movements, in which specific sites control multiple muscle groups that contribute to the generation of particular actions? Initial experiments implied that the map in the motor cortex is a fine-scale representation of individual muscles. Thus, stimulation of small regions of the map activated single muscles, suggesting that vertical columns of cells in the motor cortex were responsible for controlling the actions of particular muscles, much as columns in the somatic sensory map are thought to analyze particular types of stimulus information (see Chapter 9).More recent studies using anatomical and physiological techniques, however, have shown that the map in the motor cortex is far more complex than a columnar representation of particular muscles. Individual pyramidal tract axons are now known to terminate on sets of spinal motor neurons that innervate different muscles. This relationship is evident even for neurons in the hand representation of the motor cortex, the region that controls the most discrete, fractionated movements. Furthermore, cortical microstimulation experiments have shown that a single muscle is represented multiple times over a wide region of the motor cortex (about 2–3 mm in primates) in a complex, mosaic fashion. It seems likely that horizontal connections within the motor cortex create ensembles of neurons that coordinate the pattern of firing in the population of ventral horn cells that ultimately generate a given movement.Thus, while the somatotopic maps in the motor cortex generated by early studies are correct in their overall topography, the fine structure of the map is far more intricate. Unraveling these details of motor maps still holds the key to understanding how patterns of activity in the motor cortex generate a given movement.Figure 17.8. The corticospinal tract. Neurons in the motor cortex give rise to axons that travel through the internal capsule and coalesce on the ventral surface of the midbrain, within the cerebral peduncle. These axons continue through the pons and come to lie on the ventral surface of the medulla, giving rise to the pyramids. Most of these pyramidal fibers cross in the caudal part of the medulla to form the lateral corticospinal tract in the spinal cord. Those axons that do not cross (not illustrated) descend on the same side and form the ventral corticospinal tract (see Figure 17.6). The axons that terminate in the reticular formation of the pons and medulla comprise components of the corticobulbar tract. Transmission of Signals from the Motor Cortex to the MusclesMotor signals are transmitted directly from the cortex to the spinal cord through the corticospinal tract and indirectly through multiple accessory pathways that involve the basal ganglia, cerebellum, and various nuclei of the brain stem. In general, the direct pathways are concerned more with discrete and detailed movements, especially of the distal segments of the limbs, particularly the hands and fingers.Corticospinal (Pyramidal) TractThe most important output pathway from the motor cortex is the corticospinal tract, also called the pyramidal tract, shown in Figure 55–4.The corticospinal tract originates about 30 per cent from the primary motor cortex, 30 per cent from the premotor and supplementary motor areas, and 40 per cent from the somatosensory areas posterior to the central sulcus. After leaving the cortex, it passes through the posterior limb of the internal capsule (between the caudate nucleus and the putamen of the basal ganglia) and then downward through the brain stem, forming the pyramids of the medulla. The majority of the pyramidal fibers then cross in the lower medulla to the opposite side and descend into the lateral corticospinal tracts of the cord, finally terminating principally on the interneurons in the intermediate regions of the cord gray matter; a few terminate on sensory relay neurons in the dorsal horn, and a very few terminate directly on the anterior motor neurons that cause muscle contraction.A few of the fibers do not cross to the opposite side in the medulla but pass ipsilaterally down the cord in the ventral corticospinal tracts. Many if not most of these fibers eventually cross to the opposite side of the cord either in the neck or in the upper thoracic region.These fibers may be concerned with control of bilateral postural movements by the supplementary motor cortex.The most impressive fibers in the pyramidal tract are a population of large myelinated fibers with a mean diameter of 16 micrometers. These fibers originate from giant pyramidal cells, called Betz cells, that are found only in the primary motor cortex. The Betz cells are about 60 micrometers in diameter, and their fibers transmit nerve impulses to the spinal cord at a velocity of about 70 m/sec, the most rapid rate of transmission of any signals from the brain to the cord. There are about 34,000 of these large Betz cell fibers in each corticospinal tract. The total number of fibers in eachcorticospinal tract is more than 1 million, so these large fibers represent only 3 per cent of the total. The other 97 per cent are mainly fibers smaller than 4 micrometers in diameter that conduct background tonic signals to the motor areas of the cord.
- Corticospinal system : Role in MovementThe corticospinal and corticobulbar system is the primary pathway for the initiation of skilled voluntary movement. This does not mean that movement—even skilled movement—is impossible without it. Nonmammalian vertebrates have essentially no corticospinal and corticobulbar system, but they move with great agility. Cats and dogs stand, walk, run, and even eat if food is presented to them after complete destruction of this system. Only in primates are relatively marked deficits produced. Careful section of the pyramids producing highly selective destruction of the lateral corticospinal tract in laboratory primates produces prompt and sustained loss of the ability to grasp small objects between two fingers and to make isolated movements of the wrists. However, the animal can still use the hand in a gross fashion and can stand and walk. These deficits are consistent with loss of control of the distal musculature of the limbs, which is concerned with fine skilled movements. On the other hand, lesions of the ventral corticospinal tract produce axial muscle deficits that cause difficulty with balance, walking, and climbing. Guyton
- CEREBRAL CORTICAL CONTROL OF MOVEMENT*HIROSHI ASANUMAThe Rockefeller UniversityNew York, N.Y.Introduction:The purpose of the series of experiments which will be described in this article was to increase our understanding of how the motor cortexcontrols the movement. It has been a continuing controversy whether the motor cortex controls the contraction of single muscles or rathermuscle groups. Hughlings Jackson and his followers, based on the fact that a patient can recover the use of a limb after destruction of thatlimb area previously defined by stimulation, favored the idea of a widespread overlapping of the representation of muscle groups (cf. Ruth1965). This view was strongly supported by Sherrington’s early experiences (Hering and Sherrington, 1897, Sherrington, 1906) in whichstimulation on the surface of the cortex could not produce solitary contraction of a muscle and that the combination of the muscles activatedby cortical stimulation was always in a reciprocal fashion, i. e., “cortical stimulation never produced simultaneous contraction of the trueantagonists” (Sherrington, 1906). The latter evidence led him to think that the motor cortex controls the coordinated contraction of a groupof muscles.The interpretation that the motor cortex “thinks” in terms of movements, not muscles was so persuasive that more than half a century haspassed under the dominance of this theory. In 1947, however, Chang, Ruth and Ward demonstrated evidence that the motor cortex may be able to think in terms of muscles. They stimulated the surface of the monkey cortex, with maximum possible caution, and were able to elicit solitary contraction, although occasionally, of a single muscle. Succeeding experiments by Bernhard and Bohm (1954) demonstrated that a single shock stimulation of the motor cortex activated the fast conducting corticospinal neurons of the pyramidal tract which in turn elicited a monosynaptic response in a forelimb nerve but not in the rest of the nerves. They also showed that the cortical field which produced the monosynaptic response in a certain nerve was more restricted than the fields from which other (polysynaptic) activities in the motoneurons of the same nerve could be evoked. The study about the localization of cortical motor function was further advanced by Landgren, Phillips and Porter (1962). They stimulated the surface of the motor cortex while recording intracellularly from motoneurons and demonstrated the existante of a cluster of neurons which projected monosynaptically to motor neurons and which was confined within a narrow focus of the cortex having a cross section of the order of lmm2. Thus progress in the stimulation experiments of the motor cortex provided evidence that the contraction of individual muscles is represented in small areas of the motor cortex in a mosaic fashion. This conclusion, however, does not resolve the question of “whether the motor cortex thinks in terms of movements or muscles” because as has been pointed out by Bernhard and Bohm (1954) in their stimulation experiments, a given cortical stimulus which facilitated a monosynaptic reflex, at the same time inhibited the monosynaptic reflex of the antagonist in the absence of peripheral influences.In the meantime, another line of investigation of the motor cortex developed depending on the rapid progress of the electrical techniqueswhich enabled recording of the electrical signals associated with the activities of neurons in the central nervous system. In 1953, Malis,Pribram and Kruger showed that the motor cortex received afferent inputs from the periphery in the absence of the sensory cortex. This observation raised the question of the functional significance of these afferent inputs converging onto the motor cortex.The specific purpose of a series of experiments performed in my laboratory has been to further elucidate the details of the localizationof the cortical motor function and then to relate the motor function to the afferent inputs, i. e., to study the input-output relationships. Theterm “motor cortex” is used instead of “motorsensory cortex” because a recent histological study (Hassler and Muhs-Clement, 1964) has shownthat the cat’s pericruciate cortex corresponds to the motor cortex of the primates.
- Cellular Organization 1. The neocortex (most of the cerebral cortex) consists of six layers of histologically and functionally distinct cells2. Each region of M-1 (and the remainder of the neocortex) is organized as units of interconnected columns of several hundred neurons arranged perpendicular to the cortical surface and including all six cortical layers3. Stimulation of a given column may activate a single muscle ; more usually stimulation of a column activates several muscles to produce a coordinated movementNote: this is sometimes expressed as "The motor cortex thinks in terms of movements, not muscles"4. Groups of columns control groups of alpha motoneurons to determine the force and direction and velocity of movement 5. Descending axons forming the cortical output from M-1 (and from the cortex in general) arise from the pyramidal cells in cortical layer 56. Pyramidal cells in layers 2 and 3 send axons to other regions of the ipsilateral cortex (layer 2) and to corresponding areas of the contralateral cortex via the corpus callosum (layer 3)Note: Axial regions of the body are well represented in corpus callosal axons connecting corresponding contralateral regions of the motor cortex but distal regions are less well represented.
- Fig.1. Solid diagram of effective volume for the facilitation of monosynaptic reflexes in n. extensor carpiulnaris (ECU) within the cat’smotor cortex. Numbers indicate positions on the surface where microelectrode was inserted and heavy line show regions where intracorticalmicrostimulation (ICES) facilitated ECU monosynaptic reflexes. (Asanuma and Sakata, 1967).Fig.2. Depth - threshold relationship for direct excitation of a PT cell by intracorticalmicrostimulation (ICMS). Threshold current is plottedagainst relative depth of the stimulating electrode. Electrode depth is expressed as distance superficial or deep to the point of minimum threshold.Open circles: thresholds for cathodal pulses. Crosses: thresholds for anodal pulses. (Modified from Stoney, Thompson and Asanuma,l968).The Organization of Cortical Efferent SystemAs stated in the Introduction the short latency, low threshold effect of cortical stimulation is mediated, primarily, by the pyramidaltract (Bernhard and Bohm, 1954, Landgren et al. 1962). This tract originates from the pyramidal tract cells (PT cells) which are locatedin the deep layers of the cortex (layers V and VI) and the majority of them are located in the motor cortex. The motor area is known to bearthe thickest (up to 4.5 mm in human) cortex in the whole brain and the thickness of the motor cortex in the monkey is only slightly less thanin the human (Harman, 1947) although in the cat, it is considerably less (2.5 to 3.0 mm). To activate the PT cells from the surface, therefore,a stimulating current has to travel a long distance through the brain. This inevitably activates a large number of cortical neurons includingthe PT cells and obscures the accuracy of measuring the location of the target cells.This difficulty was overcome by stimulating the depth of the cortex directly with a microelectrode (Asanuma and Sakata 1967). Furthermoreby utilizing a closed chamber (Davies, 1956), respiratory and pulsatory movement of the brain could be minimized and the electrodecould be positioned into the depth without damaging the tissue. With this method, we found that threshold current for facilitation or inhibitionof the monosynaptic reflexes was much lower (less than l/100) than the threshold from the surface.Stimulating with weak current through the microelectrode, we found that the sites which produced facilitation or inhibition of given reflexes were confined to a narrow focus within the depth of the cortex.Figure 1 illustrates an example of the results obtained. Among several cortical penetrations six passed through a discrete region where intracorticalmicrostimulation (ICMS) of 6 pa or less facilitated monosynaptic reflexes in n. extensor carpiulnaris (wrist extensor). The effectivearea was reconstructed by histological examination and found to extend along the direction of the radial fibers within the gray matter. At theedge of the effective area, the threshold increased sharply. Other trials revealed similar results and the diameter of the effective areas, i. e.,the efferent zones, ranged from 0,5 mm to a few mm, the average being 1.0 mm.At this stage, it became imperative that the effective spread of the stimulating current within the brain be known to quantitatively analyzethe results. This was examined (Stoney, Thompson amd Asanuma 1968) by introducing 2 microelectrodes into the cortex, one stationedclose to a PT cell and the other, moved through the neighboring tissue.ICMS was delivered at various depths through the moving elect rode and the effect was monitored by recording action potentials through thestationary electrode. Figure 2 shows an example of the results. As the stimulating electrode approached the target cell, the threshold decreasedto a minimal value and then increased again. Other trials revealed similar results and in all cases, negative currents were farmore effective than positive currents and the curves always took the form of parabolas.
- Fig.4. Contractions produced in m. extensor digitorumcommunis (EDC: upper trace) and m. palmarislongus (PAL: lower trace) by intracorticalmicrostimulation (ICMS). Inset figurine shows electrode track and the foci stimulated. Stimulation at pos. 2 produced simultaneous as wellas reciprocal contraction of the 2 antagonistic muscles. The third line of each set of traces indicates ICMS. (Asanuma and Ward, 1971).Fig.5. Differentiation of excitatory area for m. pal. long.(PAL) and inhibitory area for m. ext. dig. corn. (EDC). Triangle represents thepoint which produced sustained contraction of EDC. Circles represent points which produced contraction of PAL. Stimulation of points representedby filled squares inhibited sustained contraction of EDC. Note circles and filled squares did not overlap except at one point,(Asanuma and Ward, 1971).The preceding results, indicated the existence of discrete efferent zones within the depth of the cortex which control contraction of individual muscles. Stimulation on the surface of the cortex, as stated in the introduction, never produced simultaneous contraction of the antagonists.We know, however, from our experiences that agonists and antagonists can contract simultaneously during voluntary movements. Similarily,these muscles undergo co-activation when a cat climbs a tree, hanging its body weight on the claws.To provide a solution to the question, following experiments were undertaken. Instead of monosynaptic reflexes, contraction of antagonisticmuscles, i. e., m. extensor digitorum communis (EDC, wrist flexor) and m. palmarislongus (PAL, wrist extensor) were recorded to determine the effect of ICMS. Figure 4 shows an example of the resuits.The penetration shown by the inset figurine passed through efferent zones for both EDC and PAL. When ICMS was delivered to theEDC (pos. #l) or PAL zone (pas. #3), the resulting contraction appeared in EDC or PAL, without relaxation in the antagonist as shown in Fig.4, 1 and 3. This is a striking difference from the surface stimulation.Stimulation of EDC or PAL area on the surface of the cortex under the same experimental set-ups also produced contraction of the target muscle,but in addition, that contraction was very frequently accompanied by the relaxation of the antagonist. When the intermediate zone wasstimulated, the feature of the contractions were different from when EDC or PAL zone was stimulated. As shown in Fig. 4-2, both contractionswaxed and waned, but it was observed that the contractions were not always mutually exclusive. Successive stimuli with the same intensitysometimes caused simultaneous contraction of both muscles and sometimes elicited contraction of one muscle accompanied by relaxation ofthe other. Thus when a single cortical efferent zone was stimulated, the resulting effect was a solitary contraction of the target musclewhich was not accompanied by the relaxation of the antagonist unless the intermediate zone was stimulated. We know, however, that the surface stimulation always produced contraction of the muscles in a reciprocal fashion (Sherrington, 1909). To solve the problem, two microelectrodes instead of one were inserted into the motor cortex, one into the EDC zone and the other into the PAL zone, and the interaction between the two stimuli were examined. In an example shown in Fig. 5, one electrode was positioned at an area where stimulation produced sustained contraction of EDC for the duration (1 sec. ) of the stimulus train. The effect of stimulation of PAL zone on EDC contraction was examined by delivering the second ICMS through the second electrode. Three penetrations were made in this trial and the effect of the second stimulation was checked at 0.5 mm intervals along each penetration. It was found that an area which produced contraction of PAL was located approximately 1.5 mm from the EDC point. Out of 3 sites which produced PAL contraction, only one site produced inhibition of EDC contraction. The other 2 sites produces contraction of PAL independently of EDC contraction. Furthermore, it was found that inhibition of EDC contraction was produced by stimulation of an adjacent area which did not produce contraction of PAL. In addition, in other experiment which is not shown here, stimulation which producedcontraction of EDC facilitated contraction of PAL. The results demonstrated that the area which produces contraction of a muscle is not necessarily co-extensive with the area which inhibits contraction of the antagonist. The results suggested that the corticofugal pathway which is responsible for the contraction of a muscle is different from the pathway for the inhibition of the antagonist. In other words, corticofugal fibers which activate inhibitory interneurons for a particular motoneuron pool or pools are not always the branches of the fibers which excite the motoneuron pool of the antagonist. This interpretation is also supported by the observation that the cervical interneurons which were activated by ICMS of the motor cortex were different from the interneurons activated by stimulation of group I afferent fibers (Asanuma, Stoney and Thompson, 1971). Taken together, it is suggested that the inhibitory - excitatory interaction between different motoneuron pools taking place during cortically induced movements is different from the interaction occuring during the spinal reflexes which is reciprocal in character.The difficulty in producing simultaneous contraction of agonist and antagonist by surface stimulation may be explained by the followingconsiderations. Although there is discrete spatial differentiation between the excitatory area for a given muscle and the inhibitory areafor the antagonist, these areas are located close to one another. In addition, these two areas are often located near the excitatory areafor the antagonist. Hence, current pulses applied to the surface indiscriminately activate all the regions and reveal only the predominanteffects.
- Fig.6. Distribution of effective spots within the depth of the monkey motor cortex with intracorticalmicrostimulation (ICMS) of 5 pa. Crosseson the penetrations are the spots where ICMS did not produce any movement.0: thumb flexion, 0: thumb extension, : thumb adduction,m:thumbabduction. (A sanuma and Ro&n, 1972).The columnar cortical efferent zones found in the cat’s motor cortex were also found to exist in the motor cortex of the monkey(Asanumaand Rosen, 1972). An example of the results is shown in Fig. 6 in which ICMS could produce all of four thumb movements, i. e., extension, flexion,abduction and adduction. Each efferent zone had a sharp boundary and the efferent zones controlling the various movements of a joint werelocated close together. Each efferent zone extended along the direction of radial fibers in the gray matter, as shown by the extension area inFig. 6. In addition, although not shown, it was found, by using the electromyograms as indicators, that the edge of a given efferent zone veryfrequently overlapped with the edge of another efferent zone which projected to the antagonist. This frequent overlap is a marked differencefrom the cat’s in which simultaneous contraction of the antagonists such as shown in Fig. 4 occurred only occasionally. The difference may indicatethe difference of the characteristics of the movements between these two species; the motor cortex of the monkey, especially the forelimbarea may be organized in such a way that can fix a joint by simultaneously contracting the antagonists to exert a force to the outside ofthe body whereas in the cat, it may be organized to support the body weight against the gravity (Asanuma and Rosen, 1972).
- Fig.7. Relationship between the threshold currents for direct excitation of PT cells delivered through a tungsten microelectrode and the amplitudeof the antidromic spikes of the same cells recorded by the same microelectrode. Circled points indicate overlap of values. (Stoney,Thompson and Asanuma, 1968).Afferent Inputs to Cortical Efferent Zones; Input-Output Relationships In the preceding section, it was shown that there are discrete efferentzones within the depth of the motor cortex in the cats and monkeys. A natural question then is what information do these efferentzones receive from the periphery since it is known that both the monkeys’ (Malis et al., 1953) and cats’ (Brooks Rudomin and Slayman, 1961,a, b. Buser and Imbert, 1961) motor cortices receive inputs from the periphery.Before getting into the experiments, however, some technical problems had to be solved. The basic method we used consisted in, firstrecording the unitary activities of neurons in the motor cortex to delineate their receptive fields, then stimulating the same area throughthe same micro-electrode to define the motor effects. By repeating this procedure, the relationship between afferent input to and motoroutflow from a given area of the cortex could be studied. The input-output relationships identified in this way, however, would be invalidatedif the spread of the stimulating current was wider than the extent of the area from where the electrode could record the activities of the neurons.This problem was again resolved by inserting two electrodes into the brain, one for recording unitary spikes from a PT cell and the other fordelivering ICMS. Threshold current for exciting the target PT cell was determined first by monitoring the action potentials of the target cellthrough the recording electrode. The stimulating electrode was then used as a recording electrode and the antidromic spike of the same PTcell was recorded through the same electrode. With this method, the relationship between current needed to excite a given cell from a givensite and the size of the antidromic spike of the same cell recorded at the same site could be examined. Figure 7 shows the result. When thethreshold grew higher, the spike amplitude became smaller. The majority of the neurons excited by ICMS of 10 pa or less were located closeenough to the electrode so that the activities of the same cells could be recorded by the same microelectrode. When 5 ua was used, the actionpotentials of all the cells excited could be recorded by the same electrode.Input-output relationships of discrete motor colonies were first examined in the cat by Asanuma, Stoney and Abzug (1968) and Sakataand Miyamoto (1968) and then in the monkey by Rosen and Asanuma (1972). In this section, the details about the monkey experiments willbe described because the results from both species were essentially similar, but the monkey experiments revealed greater details of therelations hips.We will start with the nature of the afferent inputs to the motor cortex. Neurons recorded during this series of experiments were classifiedinto 3 types and their distributions within the depth of the cortex are shown in Fig. 8. S-cells refers to neurons which responded to hairbending, light touch or light pressure of somewhere on the contralateral body surface. D-cells are those neurons which responded to passivejoint movements and/or pressure to deep structures. There was no difference in the depth distribution between neurons driven by superficialand deep receptors but there was a marked difference between driven cells and undriven cells. Undriven cells were found more in thedeep layers. Furthermore, cells found in the low threshold areas received more peripheral inputs than cells located in the high thresholdareas. The overall feature of the distribution of driven cells suggests that the afferent inputs from the periphery arrive in the superficiallayers of the cortex and then impulses descend synaptically to the deep layers. The fact that there were more undriven cells in the deep layersindicated that these transsynaptic impulses were not powerful enough to activate all the corticofugal neurons located in the deep layers.
- Fig.9. Reconstruction of electrode tracks and cell locations within the monkey motor cortex. Electrode penetrations (solid lines, identified bynumbers) passed through efferent zones projecting to various thumb muscles. The peripheral motor effects produced by ICMS (55 pa) are indicated by symbols explained in the figure. Cortical spots where 5 pa ICMS did not produce motor effects are shown by small solid lines perpendicular to the tracks. Positions of cells encountered are connected by dotted lines to figurines and descriptions which explain receptive fields and adequate stimuli. Spontaneously active cells not driven by peripheral stimulation are indicated as UD. (Rosen and Asanuma, 1972).An example of the distribution of S- and D- cells within a given efferent zone is shown in Fig. 9. In this experiment, the electrode was insertedinto the brain and whenever unitary activities of neurons were recorded, their receptive fields were examined and then ICMS of 5 pawas delivered. In addition, the effect of ICMS was examined at 0.5 mm intervals irrespective of the existence of the spikes. As is shown, theelectrode passed through the areas which produced flexion, extension, adduction and abduction of the thumb. The cells driven by skin stimulationshowed the characteristic localization of their receptive fields in relation to the motor effects. Two cells found in the “thumb flexioncolumn” were both activated from the ventral aspect of the thumb. The cells found in the columns for thumb extension, adduction and abductionwere activated from the distal tip, the medial aspect and the lateral aspect of the thumb respectively. The other cells found in this area wereactivated by passive thumb movements or thenar pressure although some of the cells could not be driven by natural peripheral stimulation.Other trials revealed similar results which could be summarized as follows: cells driven by peripheral stimulation were sparsely scatteredin radial columns within the hand area of the monkey motor cortex and received afferent input from superficial as well as from deep receptorslocated in restricted parts of the distal forelimb. This columnar arrangement coincided with the columnar organization of cortical efferent zonesso that each efferent column received sensory information from a portion of the limb in close anatomical relation to the muscle to which it projected.
- Fig.10. Summary of receptive fields and adequate stimuli of all driven cells encountered within efferent zones where ICMS (510 pa) producedthumb extension (A), thumb adduction (B) or thumb extension (C). The receptive fields of cells activated by tactile stimuli (S-cells) areshown in figurines on the left. The type and location of the adequate stimulus for activation of cells driven by deep receptors (D-cells) arelisted on the right. (Ros/en and Asanuma, 1972).In the following section, further details about the afferent inputs will be described. Out of 192 cells isolated from the monkey motor cortex, 37 (19%) were activated by tactile stimulation. All the receptive fields except eight were confined to non-hairy parts of the monkey’s hand. The majority of the cells adapted rapidly to maintained mechanical stimulation. Fig. 10 shows the receptive fields of all driven cells found in identifiedthumb areas. The receptive fields of all six S- cells found in thumb flexion zones were located on the ventral aspects of the thumb (Fig.10A). Similarly, all four cells located in thumb adduction zones were activated from the medial aspect of the thumb (Fig. 10B). Similar re -sults were obtained for other fingers and showed that each of the efferent zones received tactile inputs almost exclusively from that sideof the finger or hand which was in the direction of the movement produced by ICMS of the same zone.
- Inputs from deep receptors to efferent zones are summarized in Table 1. A total of 56 cells responded to stimulation of the deep structures(D-cells). Of these, 42 responded to passive joint movement and the remainder to pressure applied to deep structures. All the cells listedin the table were found at sites where muscle contraction could be produced with ICMS of 10 pa or less. Of the 42 cells activated by passivejoint movement, the majority (38 cells) were driven from the joint involved in the movement produced by ICMS. In the analysis shown inTable 1, the following assumptions were made. Cells activated by manipulation of a joint and uninfluenced by pressure applied to the bellies ofthe muscles stretched by the effective joint movement were considered to be activated by joint receptors (Table la, b). Cells driven by passive joint movement and also by pressure of the muscle stretched by the effective joint movement were considered to be activated by inputs fromthe receptors located in these muscles (Table lc). This discrimination was difficult for thumb and finger movements, but could be done easilyfor cells driven by wrist movements because of the good anatomical separation between the joint and the forearm muscle bellies. Thirteencells were found in zones projecting to wrist muscles and were driven by passive joint movements. Of these, 6 were activated by receptorsin the wrist joint. In all these cases, the direction of joint movement adequate for activating the cells was the same as that produced by ICMS.Five cells activated by a movement opposite to the direction of ICMS induced movement were activated from muscle receptors. Two cellswere driven by passive movement of the elbow. Thus the cells located in zones projecting to the wrist muscles received information from thereceptors in the wrist joint as well as the receptors in the wrist muscle to which the cortical zone projected. Cells located in the thumb andfinger zones could be driven by a passive movement opposite to that induced by ICMS as shown in Table lb, but it is possible that they wereactivated by afferent impulses from receptors situated in the intrinsic hand muscles. The muscle receptors responsible for the corticipetalimpulses are unlikely to be the annulospiral endings because group I afferent fibers have not been found to project to the motor cortex )Oscarsonand Rosen, 1963, Phillips, Powell and Wiesendanger, 1971).In summary, cells in a given cortical efferent zone receive information about the contraction of the target muscle as well as about the movementof the target joint. In addition, they receive inputs from the skin area most likely to be excited by the movement elicited by the contractionof the target muscle during the course of a manipulatory sequence. In short, the afferent inputs converge intoan efferent zone as a sequence ofthe contraction of the target muscle. What would be the functional significance of these inputs in the performance of movements? The patternof somatic sensory inputs which were found to reach the motor cortex appears to be more suited to a role as feedback information, of possibleuse for modulating the output from individual cortical columns during the movements, rather than being involved in the initiation of motor activity.The tactile placing reaction (Bard, 1938) is a reflex type movement known to be mediated by the motor cortex. Ablation of the sensory cortexabolishes the placing reaction whereas it does not abolish the evoked potential in the motor cortex (Malis, Pribram and Kruger, 1953). Automaticgrasping reaction can be elicited by tactile stimulation of the hand of monkeys and humans (Denny-Brown, 1960). This reaction does notdisappear after removal of the sensory cortex but is abolished by ablation of the motor cortex or by section of the pyramidal tract. The input-output columns described in this article may serve as the neuronal basis for this reaction although it is known that some type of grasping reaction (e. g. instinctive grasping reaction) involves sequential contraction of many muscles in a specific pattern which makes it difficult to be explained by the input-output relationships described here.
- Fig.14. Schematic diagram indicating a closed loop between the motor cortex and the periphery. Activation of a cortical efferent zone producesa contraction of the target muscle which activates receptors related to the muscle contraction and send impulses back to the originalcortical efferent zone.Final CommentsOne of the questions imposed on this series of experiments was “whether the motor cortex thinks in terms of movements or muscles?”The answer obtained was “the motor cortex thinks in terms of muscles. ” It has been shown that each muscle is represented by a small volume of the cortex which has a discrete boundary and is located within the depth of the motor cortex i. e., the cortical efferent zone. Stimulation within a given efferent zone produced contraction of a given muscle, but did not necessarily inhibit the antagonist, instead sometimes facilitated contraction of the antagonist. Studies about the afferent inputs to cortical efferent zones revealed that each efferent zone received peripheral inputs specifically related to the contraction of the muscle to which the efferent zone projected. These afferent pathways constituted ascending link of a loop circuit which connects the motor cortex and the periphery. A schematic diagram of this closed loop is shown in Fig. 14. This loop circuit by itself, however, does not explain how the movement is initiated and also how it is organized in a purposeful way. Theafferent inputs involved in this loop are topographically organized and in many cases, specific in a particular modality. These inputs are mostlikely to have been transferred through the dorsal column - medial lemniscus n. ventralisposterolateralis system as had already been inferred(cf. Brooks and Stoney, 1971). The motor cortex, however, is known to receive most powerful inputs from the n. ventralislateralis which doesnot receive inputs from the dorsal column - medial lemniscal system.Neurons in VL are shown to receive non-specific inputs from the periphery in parallel to the inputs from the cerebellum (Massion and Albe-Fessard, 1965, a, b). Stimulation of VL is known to produce powerful mono- and polysynaptic activation of PT cells (Amassian and Weiner,1966). Furthermore, it has been shown that the projection fibers from VL to the motor cortex branch somewhere in the white matter and reachdifferent parts of the motor cortex (Massion and R&pal-Padel, 1973). Obviously, the VL - Cx system has a different characteristic from thelemniscal - Cx system suggesting that the functional significance of this system is different from the loop system that we have studied. Investigation about this system has just been started in our laboratory to further increase our understanding of how the motor cortex controls the movement.
- Figure 12-3. Medial (above) and lateral (below) views of the human cerebral cortex, showing the motor cortex (Brodmann's area 4) and other areas concerned with control of voluntary movement, along with the numbers assigned to the regions by Brodmann. (Reproduced, with permission, from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) By means of stimulation experiments in patients undergoing craniotomy under local anesthesia, it has been possible to outline most of the motor projections from the motor cortex. These have been confirmed in unanesthetizedunoperated humans by PET scanning and fMRI (Figure 12-4). The various parts of the body are represented in the precentralgyrus, with the feet at the top of the gyrus and the face at the bottom (Figure 12-5). The facial area is represented bilaterally, but the rest of the representation is unilateral, the cortical motor area controlling the musculature on the opposite side of the body. The cortical representation of each body part is proportionate in size to the skill with which the part is used in fine, voluntary movement. The areas involved in speech and hand movements are especially large in the cortex; use of the pharynx, lips, and tongue to form words and of the fingers and apposable thumbs to manipulate the environment are activities in which humans are especially skilled. The conditions under which the human stimulation studies were performed precluded stimulation of the banks of the sulci and other inaccessible areas. Meticulous study has shown that in monkeys, there is a regular representation of the body, with the axial musculature and the proximal portions of the limbs represented along the anterior edge of the precentralgyrus and the distal part of the limbs along the posterior edge. Another feature of M1 is the presence of considerable overlap in the muscles that are innervated, as well as innervation of synergic muscles separated by considerable distances. There has been debate about whether individual muscles or movements are represented in M1, and the most recent evidence indicates that both are represented. The cells in the cortical motor areas are arranged in columns. The cells in each column receive fairly extensive sensory input from the peripheral area in which they produce movement, providing the basis for feedback control of movement. Some of this input may be direct, and some is relayed from somatic sensory area I in the postcentralgyrus. Cerebral dominance, which is discussed in detail in Chapter 16, also affects the motor cortex in humans. Moving the fingers of the left hand is associated mainly with activation of the right motor cortex and vice versa, as measured by imaging techniques (see Chapter 32). However, moving the fingers of the left hand also activates the left motor cortex, particularly in individuals who are right-handed. This correlates with the fact that lesions of the left motor cortex cause motor dysfunction in the left hand as well as the right hand, whereas lesions of the right motor cortex have little effect on the right hand. Supplementary Motor AreaFor the most part, the supplementary motor area projects to the motor cortex. It appears to be involved primarily in programming motor sequences. Lesions of this area in monkeys produce awkwardness in performing complex activities and difficulty with bimanual coordination. When human subjects count to themselves without speaking, the motor cortex is quiescent, but when they speak the numbers aloud as they count, blood flow increases in the motor cortex and the supplementary motor area. Thus, the supplementary motor area as well as the motor cortex is involved in voluntary movement when the movements being performed are complex and involve planning. Blood flow increases whether or not a planned movement is carried out. The increase occurs whether the movement is performed by the contralateral or the ipsilateral hand. Premotor CortexThe premotor cortex projects to the brain stem areas concerned with postural control and to the motor cortex as well as providing part of the corticospinal and corticobulbar output. Its function is still incompletely understood, but it may be concerned with setting posture at the start of a planned movement and with getting the individual ready to perform. Posterior Parietal CortexIn addition to providing fibers that run in the corticospinal and corticobulbar tracts, the somatic sensory area and related portions of the posterior parietal lobe project to the premotor area. Lesions of the somatic sensory area cause defects in motor performance that are characterized by inability to execute learned sequences of movements such as eating with a knife and fork. Some of the neurons in area 5 (Figure 12-3) are concerned with aiming the hands toward an object and manipulating it, whereas some of the neurons in area 7 are concerned with hand-eye coordination. Role in MovementThe corticospinal and corticobulbar system is the primary pathway for the initiation of skilled voluntary movement. This does not mean that movement—even skilled movement—is impossible without it. Nonmammalian vertebrates have essentially no corticospinal and corticobulbar system, but they move with great agility. Cats and dogs stand, walk, run, and even eat if food is presented to them after complete destruction of this system. Only in primates are relatively marked deficits produced. Careful section of the pyramids producing highly selective destruction of the lateral corticospinal tract in laboratory primates produces prompt and sustained loss of the ability to grasp small objects between two fingers and to make isolated movements of the wrists. However, the animal can still use the hand in a gross fashion and can stand and walk. These deficits are consistent with loss of control of the distal musculature of the limbs, which is concerned with fine skilled movements. On the other hand, lesions of the ventral corticospinal tract produce axial muscle deficits that cause difficulty with balance, walking, and climbing. Ganong 21stPrimary Motor CortexNote that more than one half of the entire primary motor cortex is concerned with controlling the muscles of the hands and the muscles of speech. Point stimulation in these hand and speech motor areas on rare occasion causes contraction of a singlemuscle; most often, stimulation contracts a group of muscles instead. To express this in another way, excitation of a single motor cortex neuron usually excites a specific movement rather than one specific muscle.To do this, it excites a “pattern” of separate muscles, each of which contributes its own direction and strength of muscle movement.Premotor AreaThe premotor area, also shown in Figure 55–1, lies 1 to 3 centimeters anterior to the primary motor cortex, extending inferiorly into the sylvian fissure and superiorly into the longitudinal fissure, where it abuts the supplementary motor area, which has functions similar to those of the premotor area. The topographical organization of the premotor cortex is roughly the same as that of the primary motor cortex, with themouth and face areas located most laterally; as one moves upward, the hand, arm, trunk, and leg areas are encountered.Nerve signals generated in the premotor area cause much more complex “patterns” of movement than the discrete patterns generated in the primary motor cortex. For instance, the pattern may be to position the shoulders and arms so that the hands are properly oriented to perform specific tasks. To achieve these results, the most anterior part of the premotor area first develops a “motor image” of the total musclemovement that is to be performed.Then, in the posterior premotor cortex, this image excites each successive pattern of muscle activity required to achieve the image.This posterior part of the premotor cortex sends its signals either directly to the primary motor cortex to excite specific muscles or, often, by way of the basal ganglia and thalamus back to the primary motor cortex. Thus, the premotor cortex, basal ganglia, thalamus,and primary motor cortex constitute a complex overall system for the control of complex patterns of coordinated muscle activity.Supplementary Motor AreaThe supplementary motor area has yet another topographical organization for the control of motor function. It lies mainly in the longitudinal fissure but extends a few centimeters onto the superior frontal cortex. Contractions elicited by stimulating this area are often bilateral rather than unilateral. For instance, stimulation frequently leads to bilateral grasping movements of both hands simultaneously; thesemovements are perhaps rudiments of the hand functions required for climbing. In general, this area functions in concert with the premotor area to provide body-wide attitudinal movements, fixation movements of the different segments of the body, positional movements of the head and eyes, and so forth, as background for the finer motor control of the arms and hands by the premotor area and primary motor cortex.
- Some Specialized Areas of Motor Control Found in the Human Motor CortexNeurosurgeons have found a few highly specialized motor regions of the human cerebral cortex (shown in Figure 55–3) that control specific motor functions. These regions have been localized either by electrical stimulation or by noting the loss of motor function when destructive lesions occur in specific cortical areas. Some of the more important regions are the following.Broca’s Area and Speech. Figure 55–3 shows a premotorarea labeled “word formation” lying immediately anterior to the primary motor cortex and immediately above the sylvian fissure. This region is called Broca’s area. Damage to it does not prevent a person from vocalizing, but it does make it impossible for the person to speak whole words rather than uncoordinated utterances or an occasional simple word such as “no” or “yes.” A closely associated cortical area also causes appropriate respiratory function, so that respiratory activation of the vocal cords can occur simultaneously with the movements of the mouth and tongueduring speech. Thus, the premotor neuronal activities related to speech are highly complex.“Voluntary” Eye Movement Field. In the premotor areaimmediately above Broca’s area is a locus for controlling voluntary eye movements. Damage to this area prevents a person from voluntarily moving the eyes toward different objects. Instead, the eyes tend to lock involuntarily onto specific objects, an effect controlled by signals from the occipital visual cortex, as explained in Chapter 51. This frontal area also controls eyelid movements such as blinking.Head Rotation Area. Slightly higher in the motor associationarea, electrical stimulation elicits head rotation. This area is closely associated with the eye movement field; it directs the head toward different objects.Area for Hand Skills. In the premotor area immediatelyanterior to the primary motor cortex for the hands and fingers is a region neurosurgeons have identified as important for “hand skills.” That is, when tumors or other lesions cause destruction in this area, hand movements become uncoordinated and nonpurposeful, a condition called motor apraxia.