1. Characterizing corticostriatal glutamatergic synaptic deterioration in HttQ111
/Q7
Huntington's Disease mouse model
1
Behavioral Neuroscience Program, Department of Psychology, Western Washington University.
*Both authors contributed equally.
Sean Andresen1*
, AJ Keefe1*
, Sydney Coffey1
, Anne Glickenhaus1
, and Jeffrey Carroll 1
Abstract
ConclusionsandDirections
Huntington's Disease (HD) is an autosomal dominant neurodegenerative
disease caused by a CAG trinucleotide expansion in the Huntingtin (Htt)
gene. The CAG trinucleotide (Q) codes for the amino acid glutamine, and
the translation of an extended polyQ sequence creates a mutant Huntingtin
protein (HTT). This protein-level mutation leads to toxic activity within the
cell, and is assumed to be the predominant cause of HD. This disease is
marked by the degeneration of various subcortical structures, particularly of
the striatum, and recent research suggests that synaptic deterioration
precedes neuronal cell loss. By comparing a mutant heterozygous
polyglutamine knock-in mouse model (HttQ111
/Q7
) of HD to a wild type
mouse model (Q7
/Q7
), we employed qualitative and quantitative methods
to characterize aberrant corticostriatal phenotypes. We compared
corticostriatal synaptic levels of VGLUT1 and PSD-95 between genotypes
using immunohistochemical and protein immunoblot methodolgies.
Aberrant levels of these proteins are classical markers for
neurodegeneration, as normal activity of these proteins is vital for neuronal
function. VGLUT1 expression is unique to corticostriatal neurons. We found
a significant loss in the glutamatergic synaptic markers VGLUT1 and PSD-95
in the mutant genotype. Additionally, astrocytic gliosis was colocalized to
areas of glutamatergic synaptic degeneration, as determined by GFAP
analysis. These results suggest a reduced quantity of corticostriatal
glutamatergic synapses in Q111/
Q7
mutant mice, potentially preceding
pathological cell loss.
Background
Huntington’s Disease (HD) is an autosomal dominant hereditary
neurodegenerative disease caused by mutations in the Huntingtin gene.
An expanded polyglutamine repeat within the gene manifests into a
disorder that destroys critical movement circuits in the brain, causes
dementia, and ultimately causes death. The subcortical grey matter
structure known as the striatum appears to be the brain region most
sensitive to HD-related neuronal degeneration. Cortical inputs to the
striatum are important for the survival of these subcortical neurons by
delivering only supply of brain derived neurotrophic factor (BDNF) into
the striatum. A protein known as TrkB acts as the postsynaptic receptor
for BDNF, and is known to be colocalized with glutamate receptors such
as AMPA and NMDA. Additionally, TrkB influences the strength with
which glutamate receptors respond to glutamate signaling. BDNF action
on postsynaptic TrkB receptors inhibit apoptosis; thus neurons not
receiving BDNF do not have this inhibitory message and undergo
apoptosis as a result. The striatum’s inability to produce its own supply of
BDNF creates a critical dependence on glutamatergic input, and a loss of
these synapses may ultimately result in neuronal death. Previous
research suggests that this may be the mechanism by which striatal
neurons undergo apoptosis in HD models. The dysfunction of synaptic
communication is a hallmark of neurodegeneration and represents an
important biomarker for disease progression.
We utilized a mutant strain of mice that are heterozygous for a mutant
htt allele that contains 111 glutamine (Q) repeats within their
homologous huntingtin gene. These mice represent an invaluable model
of HD cellular pathology. The mutant model produces a less severe and
more gradual rate of deterioration compared to other mutant mouse
strains. This allows for a more accurate representation of the human
condition and was optimal for our intentions.
Funding provided by: Huntington Society of Canada
Conclusions
Ongoing Experiments
The characterization of neural pathology in our Q111
/Q7
mouse model has provided critical endpoint biomarkers for disease
progression. We are now looking to identify abberent hepatic phenotypes that precede neural pathology. How might a
dysfunctional liver contribute to the degeneration of neurons? Do these liver cells exhibit the same abnormalities seen in
brain cells? Why are there metabolic changes that precede the degeneration of the brain, could this be causing the
disease? These questions may be centerpiece in understanding and treating the progression of Huntington's Disease. We
are in the beginning stages of developing immunohistochemical and immunoblotting techniques that can be used to
characterize hepatic phenotypes in Huntington’s Disease.
References
Blot analysis
Medium Spiny Neurons
1
BDNF
- mHtt blocks the axoplasmic transport of BDNF
leading to apoptosis in the postsynaptic neuron
VGLUT1
Apoptosis
Synapse Loss
2Gliosis
Cortex
Striatum
- Astrocytes assist in the survival of unhealthy neurons.
The presence of damaged tissue can spawn the the
inflammation of astrocytes.
- Often results in scar tissue
- Scar tissue prevents axon regrowth but effectively
seals the blood brain barrier
Corpus Callosum
- Sealing the blood brain barrier is imperative to
prevent microbes and toxins from entering the brain
Pyram
idal Neurons
3Protein Aggregation
- VGLUT1 is located presynaptically on cortical afferents. PSD-95
is located post synaptically.
PSD-95
- Huntingtin aggregates can
be found throughout an
entire neuron
- Was once believed to be
the cause of HD, but this
theory has recieved much
critisism.
- Presence of aggregates, however, is
a hallmark of HD .
- Aggregates first appear in axons and dendrites
but proteolytic activity cuts smaller fragments
that then enter the nucleus.
- They may or may not
contribute to HD pathology.
ImmunoreactiveProcess
Triton x100 is used to poke holes in cell membranes
An antibody made specifically to recognize vGLUT 1 protein
A vesicle carrying glutamate
to the synaptic button
Vglut1 Protein
Primary antibody
Secondary antibody
A secondary antibody that is conjugated
to a fluorescent molecule (FITC) is used to
detect the primary antibody
Completed complex can now be
visualized with flourecent microscopy
FITC
AXON
Antibodies are a vital tool in biological research. A“primary antibody”is used to recognize an epitope on a protein. An epitope is a
specific amino acid sequence in which the antibody binds tightly. Antibodies are actually produced inside biological organisms such
as a mouse or a rat, and extracted. An antibody that recognizes VGLUT1, for example, can be made through a process of protein
extraction and purification. First, brain tissue is removed and purified into a sample of pure VGLUT1 protein extract. The protein
sample is then injected into an organism which will then begin to produce antibodies that recognize this foreign protein circulating
its blood stream. Next the organism is sacrificed and blood serum is extracted. The antibody is purified through a process called
affinity chromatography, where the serum is passed through a matrix containing the VGLUT1 protein. Only the tissue extract that
recognizes the VGLUT1 protein (VGLUT1 antibodies) will remain in the matrix. The antibody can then be dissociated from the protein
in solution with the help of enzymes, heat, or centrifugation. This is but one method by which antibodies are manufactured. With a
price tag of roughly 350 dollars per 100 microliters, that’s equivalent to a 2 liter soda costing 700,000 dollars!
Prominent astrogliosis can be seen in this image taken from a mutant
mouse striatum. Pathologies such as synapse loss, aggregate
formation, or cell death can cause astrocytes to become inflamed. This
process serves a protective role for neuronal survival, but the resultant
scar tissue imposes complications regarding axonal regrowth. The
upregulation of astrocytes also may help to maintain the integrity of
the neural matrix as cells are lost. Post mortem HD patients show
astrogliosis that is proportional to the severity of cell loss, indicating
the importance of astrocytic analysis in our mutant mouse model.
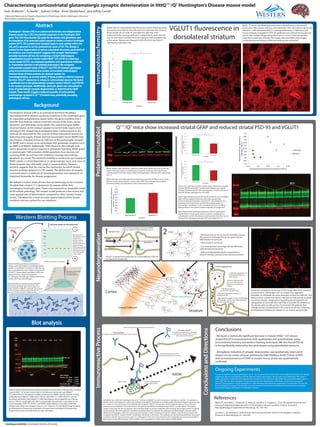
VGLUT1 fluorescence in
dorsolateral striatum
Below: 25 micron free floating sections were collected using a cryostat and
preserved in PBS solution. Sections were blocked in 10% normal horse serum and
stained in 1/400 mouse anti VGLUT1 antibody followed by 1/1000 horse anti
mouse antibody conjugated to FITC. An epifluorescent confocal microscope was
used to take multiple images along the Z-axis at .5 micron intervals and later
projected at maximum intensity. The images were quantified using ImageJ ,
unstained areas and obvious artifactual staining were subtracted.
Below: VGLUT1 immunoreactivity fluorescence levels following image
processing. Areas absent of any fluorescence represent axon tracts or
blood vessels, thus in order to normalize the data they were
subtracted before pixel quantification. A threshold for pixel intensity
was set and values recorded. Error bars represent SEM (standard error
of the mean). Mutant mice showed 34.26% (P=0.012) less VGLUT1
staining than wild type mice.
- We found a statistically significant decrease in mutant (HttQ111
/Q7
) mouse
striatal VGLUT1immunoreactivity both qualitatively and quantitatively, using
immunohistochemistry and western blotting techniques. We also found PSD-95
to be significantly reduced by the same degree using quantitative measures.
- Astrogliosis, indicative of synaptic deterioration, was qualitatively observed in
mutant mouse striata using an antibody for Glial Fibrillary Acidic Protein (GFAP),
and increased presence of GFAP in mutant mouse striata was quantitatively
confirmed.
ImmunohistochemistryProteinImmunoblotNeurodegeneration
Tissue sample (striatum)
Protein
Isolation
-
---------
------------------
---------
---------------------------
---------
---------
---------
------------------
---------
---------
---------
---------
---------
---------
---------
---------
---------
---------
------------------
---------
---------
---------
---------
---------
---------
---------
---------
---------
---------
---------
---------
---------
--------- ---------
---------
---------
---------
---------
---------
Homogenization of
protein charge
+
-
Load protein samples onto electrophoresis gel
Running current
through gel results in
movement of
proteins towards
cathode, since all
proteins possess net
negative charge.
Protein band velocity
is determined by size
(kDa): larger proteins
move through gel
matrix slowly.
Proteins transferred to membrane
Membrane is probed with
primary and then
secondary antibodies
Western blotting (protein immunoblot) is a
widely-used assay helpful in quantifying relative
protein concentrations in tissues or cells. Our aim
was to quantitatively assess relative striatal levels
of VGLUT1 (shown above and below), GFAP, and
PSD-95 between mutant heterozygous
Q111
/Q7
mouse and wild type mouse strains using
western blotting methodologies.
Primary antibodies are bound by secondary antibodies,
which possess a fluorescent tag. This tag reacts with a
certain wavelength of light (800 nm for example), and
releases a photon of a different wavelength. Spectroscopic
absorbance analysis of photon release indicates the
presence and relative amounts of labeled protein.
Western Blotting Process
FITC
Above: Western blot membrane, probed for GFAP (green bands) and actin (lower red
bands). Actin was used as a normalization control for all western blots. Membrane
contains protein from four mouse striata of each genotype: wild type (WT) and mutant
(Q111).
Below: Average normalized absorbance frequency values for all fifteen mouse striata.
Mutant mouse striata tended to contain more GFAP than wild type mouse striata,
providing evidence for astrogliosis in the mutant mouse model.
Q111
/Q7
mice show increased striatal GFAP and reduced striatal PSD-95 and VGLUT1
Above: Four wild type and four mutant mouse striata were assessed
for both PSD-95 and VGLUT1 protein levels. Mutant mice overall
showed 35.6% less VGLUT1 than wild type mice (P=0.01) and 34.7%
less PSD-95 than wild type mice (P=0.002).
Above right: Relative amounts of PSD-95 and VGLUT1 are correlated.
Both of these proteins are found in the same synapse type
(corticostriatal glutamatergic), with VGLUT1 concentrated in the
presynaptic terminal button and PSD-95 localized under the
post-synaptic membrane, so the correlated reduction in both of
these proteins in the mutant model is indicative of synapse loss.
Right: Western blot membrane probed for PSD-95 (upper green
bands), VGLUT1 (lower green bands), and actin (red bands) as a
normalization control. Membrane contains protein from four mouse
striata of each genotype: wild type (WT) and mutant (Q111).
WT Q111
WT Q111
Western blot membrane showing selected protein content of four wild type (WT) and four
mutant HttQ111
/Q7
knock-in mice (Het). In the leftmost column we used HiMark™
Pre-stained Protein Standard as a ladder, which was the ladder used for all blots. Primary
antibodies were diluted 1:2000 (actin, PSD-95, and GFAP) or 1:2500 (VGLUT1), and all
secondary antibodies were diluted 1:15000. Membranes were imaged for two minutes
each at 700 nm wavelength and 800 nm wavelength infrared light, responsible for red
(actin) and green (PSD-95, VLGUT1, and GFAP) bands (different secondary antibodies
probed actin). Blots were imaged using a Li-Cor® Odyssey® Fc Dual-Mode Imaging System,
and fluorescence frequencies of proteins of interest were normalized via fluorescence
frequencies of actin (a protein found in most cell types).
0
2
4
6
8
10
12
14
16
MeanVGLUT1fluorescenceintesityvalues
Wild Type (N=6) Mutant (N=9)
Genotype
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
Wild Type (N=8) Mutant (N=7)
AveragenormalizedGFAPabsorbance
intensity(1/M·cm)
Genotype
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0.55
0.6
0.65
0.7
0.15 0.2 0.25 0.3 0.35
NormalizedVGLUT1absorbance
intensity(1/M·cm)
Normlized PSD-95 absorbance intensity (1/M·cm)
Wild type
Mutant
Myers, R., Vonsattel, J., Paskevich, P., Kiely, D., Stevens, T., Cupples, L., et al. Decreased neuronal and
increased oligodendroglial densities in Huntington's disease caudate nucleus. Journal of
Neuropathology & Experimental Neurology, 50, 729-742.
Zuccato, C., & Cattaneo, E. Role of brain-derived neurotrophic factor in Huntington’s disease.
Progress in Neurobiology, 81, 294-330.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Wild type (N=4) Mutant (N=4)
AveragenormalizedVGLUT1absorbance
intensity(1/M·cm)
Genotype
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Wild type (N=4) Mutant (N=4)AveragenormalizedPSD-95absorbance
intensity(1/M·cm) Genotype
*
*
*