
Recommandé
Recommandé
Contenu connexe
Similaire à diss defense 1.2
Similaire à diss defense 1.2 (20)
diss defense 1.2
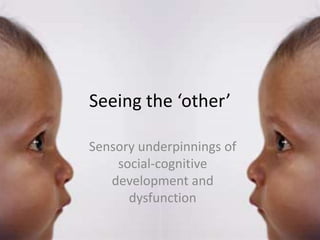
- 1. Seeing the ‘other’ Sensory underpinnings of social-cognitive development and dysfunction
- 2. Overview • Introduction • The mature neurophysiology of biological motion (BM) processing • BM processing in typical development and in ASD • BM and social cognition
- 3. Biological motion and social cognition • Social cognition - the encoding and processing in the brain of social information. • Biological motion (BM) is a prime example of a visual stimulus from which the brain extracts socially salient information.
- 5. Biological motion and social cognition • Intact BM processing is essential for healthy social interactions. • Complex social networks rely heavily on BM information to accomplish social tasks across the brain.
- 6. Mirror neuron system • “Unlike inanimate objects, humans have the distinct property of being ‘like me’ in the eyes of the observer. • This allows us to use the same systems that process knowledge about self- performed actions, self- conceived thoughts, and self- experienced emotions to understand actions, thoughts, and emotions in others.” Obermann and Ramachandran, 2007
- 7. Q: How is social information extracted from sensory data?
- 8. global STS global local “Self” percept local Auditory processing language Visual form Visual motion “Other” percept A: Not simply. EBA affect MT Motivation / Decision making motor planning cerebellum Visual input Auditory input Attentional / Motor output premotor Visual processing fusiform
- 9. Biological motion in the brain
- 10. Point-light displays • Humans are capable of detecting socially salient signals from extremely impoverished sensory information. • For example, we can detect diverse social information from so-called biological motion point light displays e.g. emotion, mood, gender, and body-type.
- 11. Signatures of biological motion • Motion coherence and opponency • Minimum jerk • Global form
- 12. Gunnar Johansson (1911-1998) Point-light displays
- 14. Biological motion • Biological motion (BM) detection is innate to the healthy human brain. • BM processing involves multiple sites across the brain. • The underlying neurophysiology remains largely unexplored.
- 15. When and where is biological motion processed in the mature brain? Is it a simple, involuntary process?
- 16. Experiment 1: Biological motion and attention neurophysiology
- 17. Biological motion and attention • Paradigm 1: Naïve subjects respond to a distractor target (dot-color) enabling the monitoring of the automatic processing of unattended BM. • Paradigm 2: Subjects are explicitly informed of the presence of BM, and respond accordingly, enabling the monitoring of attentional processes.
- 18. Biological motion and attention • High-density EEG enables a thorough mapping of bilateral visual-social pathways. • Source localization algorithms enable the estimation of neuronal generators at different temporal stages of the process. Hans Berger (1873-1941)
- 19. Initial paradigm Sample stimuli: on the left are still- frames depicting normal biological activity in point-light animation sequences. On the right are the scrambled counterparts of the biological motion sequences.
- 20. unattended attended VEPs for the unattended (a) and attended (b) biological motion (BM) tasks. Blue lines indicate the response to BM stimuli, red lines indicate the response to scrambled stimuli, and green lines represent the difference waves.
- 21. Scalp maps Posterior topographic scalp maps of the response during both experimental conditions and the difference maps between them at selected time-points.
- 22. Statistical cluster plots BM vs. SM unattended attended Color-plot of t-values for the differences between canonical biological motion point-light displays and their scrambled counterparts in the unattended (a) and attended (b) tasks.
- 23. Brain Electrical Source Analysis • BESA models intracranial dipoles that produce observed scalp waveforms, using iterative adjustments to reduce the variance between the solution and the observed data. • The upper bound of the number of modeled dipole sources was determined using an unconstrained test dipole.
- 24. Brain Electrical Source Analysis Scalp maps of the difference between BM and SM responses and the corresponding source localizations. A. Talairach: x = 35, y = -69, z = - 2; explained variance [EV] = 91% B. x = ±40, y = -69, z = 13; EV = 80% C. x = 23, y = -80, z = 20; EV = 81% D. x = ±40, y = -65, z = 7; EV = 89% E. x = -37, y = -76, z = 16; x = 32, y = -77, z = 10; EV = 93%
- 25. Brain Electrical Source Analysis Summary of findings in some recent neuroimaging studies as related to our source localizations. See, e.g., [KO] Tyler et al, 2005; [hMT] Becker et al, 2008; [pSTS] Kontaris et al, 2009. (KO=kinetic occipital area; hMT=human homolog of monkey medial temporal area; pSTS=posterior superior temporal sulcus)
- 26. How do these neurophysiological processes develop in the healthy brain?
- 27. Biological motion processing in typical development
- 28. Some basic stats Groups N (female) Age in yrs. VIQ PIQ FSIQ AS, autism cASD 35 (4) 10.48 (2.27) 7.1-15.7 101.66 (22.47) 108.26 (18.54) 105.40 (21.26) 13, 16 TDs 46 (22) 11.22 (2.90) 6.1-16.9 116.59 (14.26) 108.24 (12.88) 113.98 (13.87) N/A
- 29. New paradigm UM SM IM
- 30. Inversion
- 31. Developing biological motion VEP
- 34. TDs 10 μV PO3 PO7 P9 PO4 PO8 P10Oz Fz 100 ms unattended attended
- 35. Is biological motion processing affected in autism spectrum disorders?
- 36. Visual roots to social dysfunctions Biological motion processing in children with autism spectrum disorders
- 37. Autism spectrum disorders 1. qualitative impairment in social interaction 2. qualitative impairments in communication 3. restricted repetitive and stereotyped patterns of behavior, interests and activities
- 38. Autism spectrum disorders Select symptoms: – Has heightened or low senses of sight, hearing, touch, smell, or taste – Cannot start or maintain a social conversation – Does not adjust gaze to look at objects that others are looking at – Does not refer to self correctly (for example, says "you want water" when the child means "I want water") – Does not make friends – Does not play interactive games – May not respond to eye contact or smiles, or may avoid eye contact – May treat others as if they are objects – Prefers to spend time alone, rather than with others – Shows a lack of empathy – May withdraw from physical contact because it is overstimulating or overwhelming – Doesn't imitate the actions of others
- 39. What is the common thread?
- 40. “Mind blindness” •“extreme male brain” theory of ASD • mirror neuron system dysfunction •(for the unfamiliar?)
- 41. Affective social processing empathizing autism Ontological social processing systemizing behaviorism
- 42. ASD and biological motion Herrington et al., 2007
- 43. • There is a controversy in the literature regarding the presence of BM-processing differences in ASD. • By using high-density EEG, the spatiotemporal pathways used by TD and ASD populations can be mapped out, resolving this debate. ASD and biological motion
- 53. Mind A Interactive medium Mind B 1 0 1 0 0 1 1 1 0 Motor output Motor output Sensory input Sensory input 1 0 1 0 0 1 1 1 0 1 0 1 0 0 1 1 1 0 A A B B A B B A B A A B Representation of other’s internal states Representation of other Representation of self’s internal states Representation of self Representation of medium including BM Representation of medium including BM
- 54. External interactive environment 1 0 1 0 0 1 1 1 0 Motor output Sensory input 1 0 1 0 0 1 1 1 0 A A B B A B Representation of other’s internal states Registry of self’s internal states Representation of external environment including BM 1 1 0 1 0 1 Representation of self’s motor impact on environment Representation of other’s motor impact on environment Mind A
- 55. Conclusions • Mature BM processing: – 3 phases • Normal development: – Effects become automatic with age – No early phase – No attentional effects • ASD: – Like younger less automatic?
- 56. • Credits
- 59. Supplementary material Have some coffee first.
- 60. • Gender effects • Schizophrenia – Pilot data • The inversion effect – background • Further directions • Theory and philosophy of mind • Ncl Closure effects and “objectness” • Even more pilot analyses – Minimum norms – More clusterplots • etc.
- 63. attended ♂ ♀ ♀♂
- 64. Girls-boys minimum jerk .01
- 65. Girls-boys minimum jerk .05
- 67. Unattended – between groups ♀♂
- 69. Unattended upright-scrambled ♂; N=12 (6-12) ♀♂
- 70. TD 8-12 N=11 unattended upright-scrambled α=0.1 ♀♂
- 71. Unattended BM Gender effect 0.10 ♀♂
- 72. Unattended BM Gender effect 0.05 ♀♂
- 73. Attended BM Gender effect 0.01 ♀♂
- 74. Attended BM Gender effect 0.05 ♀♂
- 75. Attended BM Gender effect 0.10 ♀♂
- 77. Further directions Biological motion processing in schizophrenia
- 78. Schizophrenia – pilot data (N=8) unattended
- 79. Schizophrenia – pilot data (N=8) attended
- 80. Schizophrenia pilot data T-clusterplot (N=8) Unattended upright vs. scrambled Attended upright vs. scrambled
- 83. Future directions • Audiovisual eye gaze and autism (ala Klin et al, 2009) • Sensorimotor integration and ToM as a multisensory (proprioceptive) phenomenon • Electrophysiology of mentalizing and affect in ASD and/or schizophrenia • Ontological social processing… • Closure and mismatch as neurocognitive currencies • (way) future research – social-smiling infants: mindless reflex or social reward/ToM • Audiovisual interpolated and full bodies and faces vs. coherent/meaningful/inanimate motions
- 84. Low level sensory processes vs. higher order social cognition • VEPs and visual field confounds – Biological motion vs. scrambled and inverted displays – Scrambled “biological motion” vs. non-biological coherent motion (and Aristotelian spheres/ motion and mind…) – Online “minded” event and artifacts… (random structures vs. teleological structures (tools/art/etc.) – Affect and ASD
- 85. Social cognition and merging visual streams
- 86. Dorsal/ventral facial affect processing
- 87. Fusiform vs. amygdala Vuilleumier et al., 2003
- 88. Local feature and facial emotion processes
- 89. Local feature and facial emotion processes
- 90. Local feature and facial emotion processes
- 91. Higher cognitive social processing and electrophysiology • Semantic-social class formation
- 92. Social class formation • “Properties” and “objects” • Social construct formation • Theory of mind
- 93. Abstract vs. concrete perception Necessary / abstract • Conceptual • Mathematical / formal science • Possibility super-space/set • Denominator in probabilistic reasoning / stats / signal detection • E.g.: – 1+1=2 – p^~p=F Actual / concrete • Sensory • Selection • Subspace • Possibility subspace • Numerator in prob. Reasoning • E.g.: – The sky is blue. – It weighs 13 lbs.
- 94. Semantic word classes / percepts social • I • You • He • She • Person • Mind • Consciousness • Thought • See • Hear • Feel • Sense • Emotion • Want • Desire • Love • Hate • Guilt concrete • It • Inanimate • Object • Matter • Energy • Space • Time • Function • Equation • Law • Event • Particle • Motion • Position • Size • Acceleration • Color • Temperature • Sound
- 99. ToM games •Identity scales (self friend stranger foe) and BM perception in TDs and ASD
- 100. Audiovisual eye-gaze
- 101. Audiovisual Mismatch
- 102. Contextual – feedback processes
- 103. “m-eye-th”
- 105. Supplementary slides
- 106. What is “social cognition”? • Environmental processing – Sensory data (mismatches) – Data symmetries (closures) – Signal – noise inductions (closure effects) • Sense of self – Bayesian input and output – (sense of possible and actual) • Sense of other – Social signal detection
- 107. global STS global local Self percept (iostream: cognition / affect / conation) local Auditory processing language Visual form Visual motion “Other” percept Audiovisual social processes EBA affect MT Decision making motor planning cerebellum Visual input Auditory input Attentional / Motor output premotor
- 108. Animate data-sets • Perceptual orthogonality/symmetry • Objects/properties • Closure processes and mismatches • Probabilistic percepts • The “neuroeconomic” percept (“decision- maker”
- 109. (Social) signal detection • Form pathways and perceptual closure • Motion pathways and constructs – Coherence – Symmetry – Minimum jerk • Inversion effects and expert systems • Teleological processes, “mentalizing”, theory- of-mind, and beyond
- 111. Unattended
- 112. Attended
- 113. Adult results component ms task hem mot Task * hem Task * mot Hem * mot Task * hem * mot eP1 80-100 0.94 0.86 0.78 0.57 0.42 0.76 0.46 P1 100-120 0.99 0.17 0.15 0.86 0.52 0.04* 0.34 P1-N1 120-170 0.65 0.91 0.01* 0.67 0.5 0.01* 0.79 N1 170-190 0.14 0.07 0.55 0.04* 0.28 0.07 0.60 N1-P2 190-240 0.03* 0.36 0.001* 0.01* 0.01* 0.77 0.88 P2 240-320 0.04* 0.19 0.001* 0.22 0.02* 0.23 0.25 N2 320-400 0.64 0.31 0.02* 0.44 0.39 0.02* 0.18 Summary of results of 3-way ANOVA with independent variables of task-type (attended vs. unattended), hemisphere, and motion-type (BM vs. SM). (* indicates significant results at α = 0.05.)
- 115. Inversion effects - EEG Stekelenberg and de Gelder, 2004
- 116. Inversion effects - EEG Stekelenberg and de Gelder, 2004
- 117. Inversion
- 118. Inversion – biological motion Thompson et al., 2005
- 119. Inversion – biological motion Thompson et al., 2005
- 120. Inversion – biological motion Thompson et al., 2005
- 121. Social cognition in development Biological motion processing across childhood
- 122. Development Hirai et al., 2009
- 123. Development Hirai et al., 2009
- 124. Development Hirai et al., 2009 P1
- 125. Development Hirai et al., 2009 N1
- 126. Development P1-N1 Hirai et al., 2009
- 127. Development N2 Hirai et al., 2009
- 128. Development Hirai et al., 2009
- 129. TD
- 132. TD unattended upright vs. scrambled
- 133. TD attended upright vs. scrambled
- 134. Unattended upright vs. scrambled TD 6-9 yrs.
- 135. Unattended upright vs. scrambled TD 6-9 yrs.
- 136. 6-9 yrs.
- 137. Unattended upright vs. scrambled 6-9 yrs. 9-13 yrs.
- 138. Unattended RH linearity age PO8 200-300ms correlation 7.13 -2.00 -0.92 8.46^ -2.34^ 8.77 -3.87 9.79 -3.46 10.04 -5.89 10.18 -5.14 11.08 -5.37 11.89 -6.02 12.50 -6.91 ^subject is ASD sibling
- 139. Are these attended effects specific to one hemisphere?
- 140. TD attended scrambled vs. upright BM Left Right mid
- 141. TD
- 142. ASD
- 144. Both tasks collapsed
- 145. Both tasks collapsed
- 146. ASD (N=7) attended
- 147. ASD (N=7) vs. ASD (N=14) attended
- 148. PO7 ASD TD unattended attended Green=scrambled; red =upright BM; blue= Inverted BM
- 149. PO8 ASD TD unattended attended Green=scrambled; red =upright BM; blue= Inverted BM
- 150. ASD vs. TD (N=7) attended ASD TD
- 153. ASD unattended upright vs. scrambled
- 154. ASD attended upright vs. scrambled
- 155. ASD Attended upright vs. scrambled
- 156. ASD vs. TD unattended scrambled
- 157. ASD (N=14) attended upright-scrambled 180ms
- 158. However…
- 159. Unattended (N=14,14) upright-scrambled repeated measures (ordinal ages) ASD vs. TD
- 160. TD 8-12 N=11 unattended upright-scrambled α=0.1
- 161. TD 8-12 N=11 unattended upright-scrambled α=0.01, 11 consecutive timepoints
- 162. ASD attended upright-scrambled n=11; α=0.01
- 171. ASD-TD(scrambled)
- 172. ASD-TD (upright)
- 173. ASD-TD (upright)
- 175. ASD13 un
- 176. 200-300 symmetric
- 177. ASD unattended 164ms
- 179. Asd un
- 181. 395ms
- 182. TD (N=15) unattended
- 183. Td15 un upright-sc
- 184. ASD14 att
- 188. ASD vs. TD attended upright-scrambled
- 189. ASD vs. TD attended upright
- 190. ASD (N=12) vs. TD (N=10) Unattended Scrambled
- 191. ASD (n=11) vs. TD (n=10) attended scrambled
- 192. ASD vs. TD minimum jerk
- 193. 6-9.5 yrs 9.5-13 yrs. 6-13 yrs. automatic biological motion processing
- 194. closure Sehatpour et al., 2006
- 195. Sehatpour et al., 2006 closure
- 196. Weak (central) coherence • Local processing bias
- 197. • Could weak coherence early in development cause “mind blindness”?
- 198. Q: Could a simple, feed-forward sensory deficit explain social dysfunction?
- 199. A: No(?)
- 201. Mirror neuron system • “Unlike inanimate objects, humans have the distinct property of being “like me” in the eyes of the observer. • This allows us to use the same systems that process knowledge about self- performed actions, self- conceived thoughts, and self- experienced emotions to understand actions, thoughts, and emotions in others.” Obermann and Ramachandran, 2007
- 202. What’s new with mu? • Mu wave suppression and mirror neurons • Implicated in ASD? – Raymaekers et al., 2009 • No difference in ASD in contrast to other studies?
- 203. Beyond mirror neurons • 1st-person (empathetic) vs 3rd- person (systematic) processing model of neurocognition – Animacy scales – “selfness” vs. “otherness” • Extreme male brain and higher empathy/identification threshold – Reductionism: Selections and selectors • Cognition of the possible • Cognition of the actual • Non-determinism
Notes de l'éditeur
- We then conducted pointwise, paired, two-tailed t-tests between the VEP responses.
- Not seeded
- Brain electrical source analysis BESA models intracranial dipoles that produce observed scalp waveforms, using iterative adjustments to reduce the variance between the solution and the observed data. The upper bound of the number of modeled dipole sources was determined using an unconstrained test dipole. a. Scalp map of the difference between BM and SM responses at ~140 ms in the unattended task and the corresponding source localization for the 120-160 ms time-window (Talairach: x = 35, y = -69, z = -2; explained variance [EV] = 91%). b. Scalp map of the difference between BM and SM responses at ~275 ms in the unattended task and the corresponding symmetric sources localized for the 200-350 ms time-window (Talairach: x = ±40, y = -69, z = 13; EV = 80%). c. Scalp map at ~140 ms of difference-waves between scrambled and canonical biological motion for the attended task and the corresponding source localized for the 120-160 ms time-window (Talairach: x = 23, y = -80, z = 20; EV = 81%). d. Scalp map at ~275 ms of the difference between scrambled and canonical biological motion for the attended task and the corresponding symmetric sources localized for the 200-350 ms time-window (Talairach: x = ±40, y = -65, z = 7; EV = 89%). e. Scalp map at ~450 ms of the difference between scrambled and canonical biological motion for the attended task and the corresponding sources localized for the 400-500 ms time-window (Talairach: x = -37, y = -76, z = 16; x = 32, y = -77, z = 10; EV = 93%).
- Brain electrical source analysis BESA models intracranial dipoles that produce observed scalp waveforms, using iterative adjustments to reduce the variance between the solution and the observed data. The upper bound of the number of modeled dipole sources was determined using an unconstrained test dipole.
- Systemizers vs. empathizers Uta frith Simon baron cohen
- Despite similar behavioural performance, significantly less activity was found for the AS group (relative to a control group) in inferior, middle and superior temporal regions, including the human analogue of MT+/V5. These data suggest that AS is associated with unique patterns of brain activity during the perception of visually presented social cues. Areas of reduced brain activity among individuals with Asperger Syndrome during the randomised and nonrandomized walker conditions. The statistical maps represent the comparison of brain activity across the two groups for the contrast of non-randomised walker versus fixation (top) and randomised walker vs. fixation (bottom), overlayed on a template brain that has been normalised into the MNI coordinate systems and rendered using mri3dX (http://www.jiscmail.ac.uk/lists/mri3dx.html). Clusters are significant at p < .004 for the nonrandomized walker contrast, and .006 for the randomised walker contrast (each corresponding to a Type I error probability of less than one cluster). Activations in yellow represent clusters of activity that significantly lower for the AS group as compared to controls.
- Unattended upright-scrambled ♂; N=12 (6-12); Attended upright-scrambled ♂; N=12 (6-12); Attended upright-scrambled ♀; N=6 (6-11 years)
- Faces with a normal (intact) broad spatial frequency (BSF) content (left column) were filtered to contain only a high range or low range of spatial frequencies (HSF or SLF; middle and right columns respectively). Six possible face types were equally distributed in a 2 3 factorial design (n = 40 per cell), with either a fearful or neutral expression (half each), and either a BSF, LSF or HSF content (all shown in random order; duration 200 ms each; mean interstimulus interval 6.1 s). Individual face stimuli were counterbalanced across participants, with different expressions and spatial frequency contents chosen for each face across participants. In addition, each individual was repeated once (after a randomly chosen lag of between 25 and 35 intervening stimuli) with either the same or a different spatial frequency content (that is, same or different image of the same person; n = 20 faces for each type of repetition; see Results). High and low spatial frequency information in visual images is processed by distinct neural channels. Using event-related functional magnetic resonance imaging (fMRI) in humans, we show dissociable roles of such visual channels for processing faces and emotional fearful expressions. Neural responses in fusiform cortex, and effects of repeating the same face identity upon fusiform activity, were greater with intact or high-spatial-frequency face stimuli than with low-frequency faces, regardless of emotional expression. In contrast, amygdala responses to fearful expressions were greater for intact or low-frequency faces than for high-frequency faces. An activation of pulvinar and superior colliculus by fearful expressions occurred specifically with low-frequency faces, suggesting that these subcortical pathways may provide coarse fear-related inputs to the amygdala.
- History of motion capture - late 1800s - scientist Étienne-Jules Marey and Eadweard Muybridge conducted independent studies of human and animal motion. used multiple photographs of moving objects to present the biological movement (Menache, 2000). major impact in biology, medicine, photography and nowadays also in animation (Menache, 2000).
- VEPs for the unattended BM task. Blue lines indicate the response to BM stimuli, red lines the response to scrambled stimuli, and green lines represent the difference waves.
- VEPs for the attended BM task. Blue lines indicate the response to BM stimuli, red lines the response to scrambled stimuli, and green lines represent the difference waves.
- Stimulus conditions. A, Experiment 1: intact unoccluded, intact occluded, apart unoccluded, and apart occluded stimuli. B, Experiment 2: upright unoccluded, upright occluded, inverted unoccluded, and inverted occluded stimuli. C, Examples of Y (anteroposterior) and Z (inferior–superior) rotation of one gait from two parent segments and their children as a function of video frame (parent, left arm; children, left forearm, left hand; parent, right leg; children, right thigh, right shin). Note that the same Y, Z (and X, not shown) rotation for each segment was used in the intact and apart conditions. D, Cross-correlations of Y and Z rotation from the left hand and left forearm (blue), right thigh (red), and right shin (green) Y rotation as a function of frame. The cross-correlation between Y (or X or Z) rotations of different segments was identical across intact and apart conditions. Expt, Experiment.
- Overlap in effects between experiments 1 and 2. Group analysis activation data overlaid on a single subject’s normalized SPGR images (top) and inflated Colin brain atlas anatomical images of the left (bottom left) and right (bottom right) lateral hemispheres. Green indicates significant activation to all experimental conditions from experiments 1 or 2 versus fixation. Red indicates a significantly greater response to intact versus apart, regardless of occlusion from experiment 1. Yellow indicates interaction between intactness and occlusion, from experiment 1. Purple indicates a significantly greater response to upright versus inverted, regardless of occlusion, from experiment 2. Symbols indicate location of peak activation in EBA in previous studies. STG, Superior temporal gyrus; pITS, inferior temporal sulcus, posterior segment; L, left; R, right.
- Experiments (Expts) 1 and 2: mean1 SE percentageMRsignal change (% signal change) relative to fixation baseline at maximally responding anatomical locations (with corresponding centroids in Talairach coordinates). Data from experiments 1 and 2 are presented in left and right columns, respectively. White bars represent intact (Expt 1) or upright (Expt 2) conditions, gray-filled bars represent apart (Expt 1) or inverted (Expt 2) conditions. The activations for the right STS and right ITS were presented originally in Figure 2, left POF in Figure 3, and right STS/STG and right pITS in Figure 4. Unoccl, Unoccluded; Occl, occluded; POF, parietooccipital fissure; STG, superior temporal gyrus; pITS, inferior temporal sulcus, posterior segment.
- The averaged amplitude and latency of the P1 component. (A) O1/O2 electrodes and (B) T5=/T6= electrodes. The error bars indicate the standard errors (SE) of the mean.
- The averaged amplitude and latency of the N1 component. (A) O1/O2 electrodes and (B) T5=/T6= electrodes. The error bars indicate the standard errors (SE) of the mean.
- The averaged P1-N1 amplitude. (A) O1/O2 electrodes and (B) T5=/T6= electrodes. The error bars indicate the standard error (SE) of the mean.
- The averaged amplitude and latency of the N2 component at the T5=/T6= electrodes. The error bars indicate the standard errors (SE) of the mean.
- Regression analysis of the P1 amplitude (left panel) and N1 latency (right panel) at the (A) O1/O2 and (B) T5=/T6= electrodes. We found a significant negative correlation except for the N1 latency of the sPLW stimulus at T5= electrode (see Table 3).
- Dark=attended; light=unattended; green=scrambled; red=upright BM; blue=inverted BM
- Dark=attended; light=unattended; green=scrambled; red=upright BM; blue=inverted BM
- Dark=attended; light=unattended; green=scrambled; red=upright BM; blue=inverted BM
- Black=TD scrambled; red=TD upright BM; blue=ASD scrambled; pink = ASD upright
- Black=TD scrambled; red=TD upright BM; blue=ASD scrambled; pink = ASD upright
- 7 subjects who may have been actually doing task. (more precisely, 5 clearly were) 2?
- Left hem and N300
- N=10 subj
- Room for a sub-diagnostic?
- Anterior-posterior for power?
- By hemisphere
- LOC and Ncl
- Systemizers vs. empathizers Uta frith Simon baron cohen