Fiziologija bubrega
•Télécharger en tant que DOC, PDF•
2 j'aime•2,283 vues

Fiziologija bubrega

Recommandé
Contenu connexe
Tendances
Tendances (20)
En vedette
En vedette (16)
Similaire à Fiziologija bubrega
Similaire à Fiziologija bubrega (20)
Plus de Anatomija dr Šarac
Plus de Anatomija dr Šarac (20)
Fiziologija bubrega
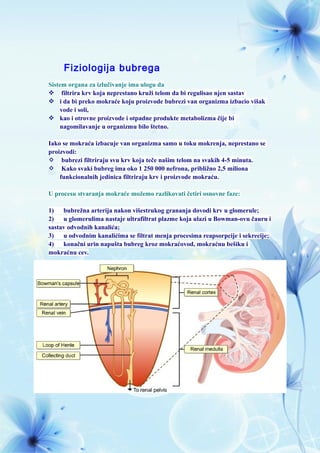
- 1. Fiziologija bubrega Sistem organa za izlučivanje ima ulogu da filtrira krv koja neprestano kruži telom da bi regulisao njen sastav i da bi preko mokraće koju proizvode bubrezi van organizma izbacio višak vode i soli, kao i otrovne proizvode i otpadne produkte metabolizma čije bi nagomilavanje u organizmu bilo štetno. Iako se mokraća izbacuje van organizma samo u toku mokrenja, neprestano se proizvodi: bubrezi filtriraju svu krv koja teče našim telom na svakih 4-5 minuta. Kako svaki bubreg ima oko 1 250 000 nefrona, približno 2,5 miliona funkcionalnih jedinica filtriraju krv i proizvode mokraću. U procesu stvaranja mokraće možemo razlikovati četiri osnovne faze: 1) bubrežna arterija nakon višestrukog grananja dovodi krv u glomerule; 2) u glomerulima nastaje ultrafiltrat plazme koja ulazi u Bowman-ovu čauru i sastav odvodnih kanalića; 3) u odvodnim kanalićima se filtrat menja procesima reapsorpcije i sekrecije; 4) konačni urin napušta bubreg kroz mokraćovod, mokraćnu bešiku i mokraćnu cev.
- 2. Sistem cevčica u svakom nefronu u proseku je dugačak oko 3cm i širok 0,05mm. Kada bi se raspleli svi tubulusi bubrega njihova dužina bi iznosila oko 30km. Dok glomerulski filtrat teče kroz tubule, 99% vode i varijabilne količine rastvorenih materija se normalno reapsorbuju, a mala količina nekih materija se šećernira u tubule. Ono što na kraju izađe iz tubula je urin. Nefron čisti krvnu plazmu od nepoželjnih materija tako što se veliki deo plazme otfiltrira kroz glomerulsku membranu u tubule. Dok otfiltrirana tečnost prolazi kroz kanaliće, nepoželjne materije se ne apsorbuju, a poželjne, posebno voda i većina elekrolita se reapsorbuju kroz plazmu koja protiče pretubularnim kanalićima. Renalna frakcija je onaj deo ukupnog minutnog volumena srca koji protiče kroz bubrege. Izražava se u procentima i kod ljudi ima vrednost od 0,12 (12%) do 0,30 (30%). Količina izlučivanja pojedinih supstanci mokraćom određena je sa tri bubrežna procesa: 1) glomerularnom filtracijom; 2) reapsorpcijom određenih supstanci iz bubrežnih kanalića u krv; 3) sekrecijom supstanci iz krvi u bubrežne kanaliće. Stvaranje mokraće započinj e filtracijom velikih količina tečnosti iz glomerularnih kapilara u Bowman-ovu čauru. Odnosno, tečnost koja nastaje filtracijom u glomerulu se naziva glomerulski filtrat i ona se uliva u mokraćni prostor između dva sloja Bowman-ove čaure. Većina supstanci iz krvne plazme, osim proteina, slobodno se filtrira, pa je njihova koncentracija u glomerularnom filtratu Bowman-ove čaure gotovo jednaka kao i u krvnoj plazmi, tj. glomerularni filtrat je izotoničan sa krvnom plazmom. Razlikuju se po tome što u glomerularnom filtratu nema eritrocita,
- 3. a proteina ima manje od 0,3g/l, pa se ovakav ultrailtrat naziva još i primarna mokraća. Na poprečnom preseku građe glomerularne membrane, tj. membrane glomerulskih kapilara, mogu se uočiti tri sloja: 1) endotel kapilara; 2) bazalna membrana; 3) sloj epitelnih ćelija koje oblažu spoljašnju površinu glomerulskih kapilara. Kapilarne endotelne ćelije su perforirane hiljadama rupica koje se nazivaju fenestre. Drugi sloj je propustljiva bazalna membrana, dok treći sloj ove membrane ne čini kontinuirani sloj. On je izgrađen od prstiju epitelnih ćelija, koji omeđavaju proreze nazvane „pukotinaste pore“. Kroz ove slojeve filtriraju se tečnost i rastvorene supstance, izuzimajući proteine i ćelije krvi. Glomerulska kapilarna membrana je permeabilna u odnosu na membrane kapilara inače. Kroz glomerularnu membranu se filtrira nekoliko stotina puta više vode i rastvorenih supstanci nego kroz obične kapilarne membrane. Kada filtrirana tečnost proteče kroz kanaliće, sastav joj se menja zbog reapsorpcije vode i određenih supstanci u krv, ali i zbog sekrecije određenih supstanci iz peritubularnih kapilara u kanaliće. Sastav mokraće određuju: 1) ultrastrukture glomerularnih kapilara; 2) poprečni presek kroz glomerularnu membranu. Veličinu glomerularne filtracije određuju: 1) hidrostatičke i koloidno-osmotske sile koje deluju na glomerularnu membranu (neto-filtracijski pritisak); 2) filtracijski koeficijent glomerularnih kapilara. Neto-filtracijski pritisak uzrokuju: 1) hidrostatički pritisak unutar glomerularnih kapilara; 2) hidrostatički pritisak u Bowman-ovoj čauri; 3) koloidno-osmotski pritisak plazmatskih proteina u glomerularnim kapilarima; 4) koloidno-osmotski pritisak proteina u Bowman-ovoj čauri. Na koloidno-osmotski pritisak u glomerularnim kapilarima utiču:
- 4. 1) koloidno-osmotski pritisak arterijske plazme; 2) udeo plazme koji se filtrira kroz glomerularne kapilare (filtracijska frakcija). Sa povišenjem, odnosno sniženjem glomerularnog hidrostatičkog pritiska povećava se, odnosno smanjuje minutna glomerularna filtracija. Veliki bubrežni protok krvi obezbeđuje dovoljno plazme za održavanje velike minutne glomerularne filtracije koja je potrebna za nadzor nad volumenom telesnih tečnosti, nad koncentracijom njihovih sastojaka. Prilikom promena bubrežnog arterijskog pritiska postoji autoregulacija bubrežnog protoka krvi i minutne glomerularne filtracije. Proksimalni kanalići reapsorbuju
- 5. oko 65% filtriranog natrijuma, hlorida, hidrogenkarbonata i kalijuma, i praktično svu glukozu, aminokiseline (glukoza ostaje samo u tragovima, i to 0,01-0,02%), kao i vitamine. Za apsorpciju glukoze, aminokiselina i vitamina je neophodna energija, jer se vraćaju u krv aktivnim transportom, procesom pinocitoze. a u prvoj polovini proksimalnog kanalića reapsorbuje se natrijum u kotransportu sa glukozom, aminokiselinama i drugim rastvorenim supstancama. U drugoj polovini natrijum se reapsorbuje zajedno sa hloridnim jonima. Proksimalni kanalići šećerniraju u tubularni lumen organske kiseline i baze, poput žučnih soli, oksalata, urata i kateholamina, kao i vodonikove jone. Silazni krak tankog segmenta Henle-ove petlje je veoma propustan za vodu, a umereno propustan za većinu rastvorenih supstanci, uključujući ureu i natrijum. Približno 20% filtrirane vode reapsorbuje se u tankom silaznom kraku. Tanki i debeli deo uzlaznog kraka skoro su nepropusni za vodu, što je važno za stvaranje koncentrovane mokraće. Debeli segment Henle-ove petlje reapsorbuje oko 25% filtriranog natrijuma, hlorida i kalijuma, znatne količine kalcijuma, hidrogenkarbonata i magnezijuma. Ovaj segment šećernira vodonikove jone . Reapsorpcijska moć tankog segmenta uzlaznog kraka mnogo je manja od reapsorpcijske moći debelog segmenta. U distalnom kanaliću se vrši reapsorpcija vode i elektrolita, Početni distalni kanalić reapsorbuje natrijum, hloride, kalcijum i magnezijum, a uopšte ne propušta vodu i ureu. Završni distalni kanalić i kortikalnu sabirnu cev izgrađuju dve različite vrste epitelnih ćelija, glavne i umetnute. Glavne reapsorbuju natrijumove jone iz lumena i šećerniraju kalijumove jone u lumen. Umetnute reapsorbuju kalijumove i hidrogenkarbonatne jone iz lumena i šećerniraju vodonikove jone u lumen. Reapsorpcija vode iz ovog segmenta je pod nadzorom antidiuretskog hormona ili vazopresina (ADH). Ovo predstavlja takozvanu nervno-humoralnu kontrolu. Naime, vazopresin, koji se stvara u hipotalamusu, a skladišti u neurohipofizi, oslobađa se na stimulaciju osmoreceptora, i pri njegovoj visokoj vrednosti tubularni segmenti su propusni za vodu i dolazi do inhibicije retencije natrijuma. Kada nema antidiuretskog hormona, oni uopšte ne propuštaju vodu. Na taj način se nadzire stupanj razređivanja ili koncentrovanja mokraće. Medularne sabirne cevi aktivno reapsorbuju natrijum i aktivno šećerniraju vodonikove jone. Unutrašnja medularna sabirna cev propusna je za ureu koja se reapsorbuje. Reapsorpciju vode iz medularnih sabirnih cevi nadzire ADH.
- 6. Neto-reapsorpcijska sila skup je hidrostatičkih i koloidno-osmotskih sila koje pogoduju reapsorpciji kroz zid peritubularnih kanala ili joj se opiru. Te sile obuhvataju: 1) hidrostatički pritisak unutar peritubularnih kapilara koji se opire reasorpciji; 2) hidrostatički pritisak u bubrežnom intersticijumu koji pogoduje reasorpciji; 3) koloidno-osmotski pritisak plazmatskih proteina u peritubularnim kapilarima koji pogoduje reasorpciji; 4) koloidno-osmotski pritisak proteina u bubrežnom intersticijumu koji se opire reasorpciji. Sile koje povećavaju reasorpciju u peritubularne kapilare, povećavaju i reasorpciju iz bubrežnih kanalića. Hemodinamičke promene koje koče reasorpciju u peritubularne kapilare, koče i tubularnu reasorpciju vode i rastvorenih supstanci. Aldosteron povećava reasorpciju natrijuma i sekreciju kalijuma. Naročito deluje na glavne epitelne ćelije završnog distalnog kanalića i sabirne cevi. Angiotenzin II povećava reasorpciju natrijuma i vode, on pospešuje lučenje aldosterona, sužava eferentne arteriole. Takođe, jednako pospešuje reasorpciju natrijuma u proksimalnim kanalićima, Henle-ovom petljama i distalnim kanalićima. ADH povećava reasorpciju vode povećavajući propusnost završnog distalnog kanalića i sabirne cevi, tako da kada nema ADH bubrezi izlučuju velike količine razređene mokraće. Aktivacija simpatičkog nervnog sastava povećava reasorpciju natrijuma. Kad je osmolarnost telesne tečnosti veća od normalne vrednosti, zadnji režanj hipofize luči više ADH, i time omogućava reasorpciju velikih količina vode što dovodi do smanjenja volumena mokraće, a da se pritom znatnije ne promeni brzina izlučivanja rastvorenih supstanci. Kada je u telu smanjena osmolarnost telesne tečnosti, smanjuje se lučenje ADH i time se ujedno smanjuje propusnost za vodu u završnim distalnim kanalićima i u sabirnim cevima, zbog čega se izlučuju velike količine razređene mokraće. Tečnost koja napušta Henle-ovu petlju je razređena, ali postaje koncentrovana kada se voda reapsorbuje iz završnog distalnog kanalića i sabirne cevi. Kada je vrednost ADH visoka, osmolarnost mokraće je približno jednaka osmolarnosti u bubrežnom medularnom intersticijumu. Urea, koja se iz unutrašnje medularne sabirne cevi reapsorbuje u intersticijumovu tečnost, ulazi u Henle-ovu petlju, a zatim se kroz distalni kanalić, kortikalnu i spoljašnju medularnu sabirnu cev vraća u unutrašnju medularnu sabirnu cev. Recirkulacija uree omogućava da se urea zarobi u bubrežnoj srži, čime doprinosi hiperosmolarnosti bubrežne srži.
- 7. Sekundarna mokraća, kao finalna mokraća, nastala delovanjem mehanizama za koncentrovanje i razređivanje mokraće, izlučuje se mehanizmom koji se ogleda u povećavanju propustljivosti epitelnih ćelija distalnog i sabirnog dela kanalića za vodu. Pod uticajem vazopresina, kada razređena tečnost iz uzlaznog kraka Henle-ove petlje pređe u distalni i sabirni deo, voda difunduje u pretubularnu tečnost, prema višem osmolaritetu. Joni sulfata, fosfata i nitrata reapsorbuju se kao joni urata. Pri reasorpciji fosfata važnu ulogu ima i hormon paratiroidne žlezde, kao inhibitor apsorpcije . Kreatinin, urea, urati (soli nokraćne kiseline) i drugi produkti katabolizma azotnih jedinjenja se uglavnom ne apsorbuju. Povratna sprega bubrezi – telesna tečnost deluje tako da sprečava trajno skupljanje soli i vode u telu pri njihovom povećanom unosu. Sve dok je funkcija bubrega normalna, velike promene unosa soli i vode uzrokovaće samo male promene volumena krvi, srčanog minutnog volumena i arterijskog pritiska. U slučaju kada je unos tečnosti manji od normalnog, postoji mogućnost smanjenja volumena krvi i snižavanja arterijskog pritiska. Čak i malo sniženje krvnog pritiska uzrokuje veliko smanjenje izlučivanja mokraće, a to omogućava održavanje ravnoteže tečnosti uz minimalne promene krvnog pritiska i volumena krvi. Održavanje acido-bazne ravnoteže – stalna koncentracija vodonikovih jona omogućena je time što bubrezi izlučuju mokraću pH vrednosti od 4,5 do 8,5. To predstavlja deo puferskog sistema organizma, koji je omogućen sekrecijom vodonikovih jona koja se odvija duž kanalića nefrona, sekrecijom amonijaka, sintezom ugljene kiseline, reapsorpcijom natrijuma. Prema tome, vodonikovi joni se neutrališu bikarbonatnim, fosfatnim i amonijačnim puferskim sistemima. Sposobnost neutralizacije toksičnih jedinjenja, kao što su fenoli, koji se u jetri i bubrezima vezuju sa sumpornom kiselinom pri čemu grade estre, i kao takvi bivaju izlučeni. Bubrezi, takođe, uzimajući aminokiseline iz krvne plazme, koriste ih za sintezu specifičnih proteina.