Transcriptomics: A Tool for Plant Disease Management
Erickson Presentation
1. I. Abstract
The
evolu)onary
mechanism
of
gene
evolu)on
has
been
studied
in
many
organisms,
but
there
is
s)ll
a
great
deal
unknown
about
this
process.
To
be<er
comprehend
this
mechanism,
we
used
Agrobacterium
as
a
model
organism.
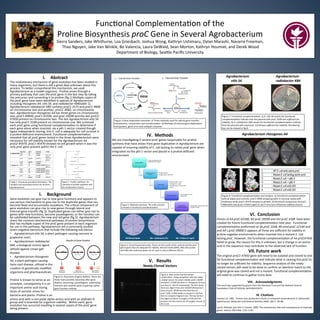
Proline
arises
through
a
primary
pathway
that
uses
the
proC
gene
in
the
last
step
by
taking
the
precursor
and
conver)ng
it
to
proline
(fig
1)
Mul)ple
copies
of
the
proC
gene
have
been
iden)fied
in
species
of
Agrobacterium
including
rhizogenes
A4,
vi7s
S4,
and
radiobacter
K84(table
1).
Agrobacterium
rabiobacter
K84
contains
proC1-‐3173
and
proC1-‐9000
on
chromosome
one
and
another,
proC2-‐
8082,
on
chromosome
two.
Agrobacterium
rhizogenes
A4
has
three
genes
on
chromosome
one,
proC1-‐99000,
proC1-‐65500,
and
proC-‐39290
and
the
last
proC2-‐
47450
present
on
chromosome
two.
The
last
Agrobacterium
vi7s
S4
has
only
proC1-‐3168
present
on
chromosome
one.
We
screened
each
proC
gene
using
func)onal
complementa)on
to
determine
if
each
gene
alone
when
inserted,
via
a
pKt-‐1
vector
created
though
ligase
independent
cloning,
into
E.
coli
is
adequate
for
cell
survival
in
a
proline-‐deficient
environment.
Func)onal
complementa)on
revealed
that
all
proC
genes
tested
in
the
three
Agrobacterium
were
necessary
for
cell
viability
except
for
the
Agrobacterium
A4
proC2-‐45470.
proC1-‐45470
showed
no
cell
growth
when
it
was
the
only
proC
gene
present
within
the
E.
coli.
I. Background
Gene
evolu)on
can
give
rise
to
new
gene
func)ons
and
appears
to
use
various
mechanisms
to
give
rise
to
the
duplicate
genes
that
can
become
fixed
and
accumulate
muta)ons.
The
cri)cal
component
of
gene
evolu)on
can
give
rise
to
new
genes
through
lateral
and
internal
gene
transfer
(fig
3).
Duplicated
genes
can
either
give
rise
to
genes
with
new
func)ons,
become
psuedogenes,
or
the
func)on
can
be
subdivided
between
the
new
and
old
gene
(fig
2).
Agrobacterium
share
the
common
biochemical
pathways
of
proline
biosynthesis
that
has
mul)ple
copies
of
the
proC
gene
present
in
each
organism
for
use
in
this
pathway.
Agrobacterium
are
a
commonly
studied
Gram-‐nega)ve
bacterium
that
include
the
following
sub-‐Genus:
• Agrobacterium
vi7s
S4,
a
plant
pathogen
causing
necrosis
in
grapevine
roots.
• Agrobacterium
radiobacter
K84,
a
biological
control
agent
u)lized
against
crown
gall
disease.
• Agrobacterium
rhizogenes
A4,
a
plant
pathogen
causing
hairy
root
disease,
u)lized
in
the
crea)on
of
gene)cally
modified
organisms
and
pharmaceu)cals.
Proline
is
known
to
serve
as
an
osmolyte,
consequently
it
is
an
important
amino
acid
during
)mes
of
osmo)c
stress
for
bacteria
and
plants.
Proline
is
an
amino
acid
with
a
non-‐polar
alpha-‐amino
acid
with
an
alipha)c
R-‐
group
and
is
essen)al
for
organism
viability.
Within
each,
gene
evolu)on
has
occurred
resul)ng
in
several
copies
of
the
proC
gene
being
present.
Func)onal
Complementa)on
of
the
Proline
Biosynthesis
proC
Gene
in
Several
Agrobacterium
Sierra
Sanders,
Jake
Whithurse,
Loa
Griesbach,
Joshua
Wong,
Kathryn
Ushimaru,
Dylan
Marashi,
Navarre
Freeman,
Thao
Nguyen,
Jake
Van
Winkle,
Bo
Valencia,
Laura
DeWald,
Sean
Morton,
Kathryn
Houmiel,
and
Derek
Wood
Department
of
Biology,
Sea<le
Pacific
University
IV. Methods
We
are
inves)ga)ng
if
several
proC
genes
responsible
for
proline
synthesis
that
have
arisen
from
gene
duplica)on
in
Agrobacterium
are
capable
of
ensuring
viability
of
E.
coli
lacking
its
na)ve
proC
gene
when
integrated
via
the
pKt-‐1
vector
and
placed
in
a
proline
deficient
environment
V. Results
Newly
Cloned
Vectors
Isolate gDNA
• Using an extraction kit
isolate all ProC genes
Amplify gene of
interest
• Design primers
• Amplify gene with PCR
Clone gene
• Preparing pKt vector
• Treatment of vector
• Hybridization of insert and
vector
Sequence gene
• Send sequence to be
sequenced
Functional
complementation
• Streak plates with culture and
compare results
Agrobacterium
Agrobacterium
vi/s
S4
radiobacter
K84
Agrobacterium
rhizogenes
A4
VI. Conclusion
Clones
of
A4
proC-‐65500,
A4
proC-‐39290
and
AVI
proC-‐3168
have
been
created
for
future
func)onal
complementa)on
next
year.
Func)onal
complementa)on
preformed
on
S4
proC-‐3168,
K8
arad
proC-‐12144
and
and
A4’s
proC-‐99000
it
appears
all
three
are
sufficient
for
viability
in
proline
nega)ve
environments
when
inserted
into
a
mutant
E.
coli
lacking
proC.
However,
the
func)onal
complementa)on
of
A4
proC47450
failed
to
grow,
the
reason
for
this
is
unknown,
but
a
change
in
an
amino
acid
in
the
sequence
may
contribute
to
the
observed
lack
of
func)on.
VII. Future
work
The
original
proC1-‐47450
gene
will
need
to
be
isolated
and
cloned
to
test
for
func)onal
complementa)on
and
indicate
what
is
causing
this
proC
to
no
longer
be
sufficient
for
viability.
Sequence
analysis
of
the
newly
cloned
vectors
will
need
to
be
done
to
confirm
an
iden)cal
match
to
the
original
gene
was
cloned
and
not
a
mutant.
Func)onal
complementa)on
will
need
to
con)nue
to
gather
more
data.
VIII. Acknowledgments
This
work
was
supported
by
grants
from
the
Murdock
Trust
and
the
Na)onal
Science
founda)on
(TUES
#7325436,
MCB
#0523357).
IX. Cita@ons
Csonka
L.N.
1981.
Proline
over-‐produc)on
results
in
enhanced
osmotolerance
in
Salmonella
typhimurium.
Molecular
and
General
Gene7cs
MGG.
182:1.
82-‐86.
Lynch,
Michael,
and
John
S.
Conery.
2000.
The
evolu)onary
fate
and
consequences
of
duplicate
genes.
Science
290.5494:
1151-‐1155.
Table
1.
Previously
provided
informa)on.
The
loca)on
and
associated
vectors
for
all
three
Agrobacterium.
Figure
3
Gene
duplica)on
overview.
a)
Three
methods
used
for
lateral
gene
transfer;
transduc)on,
conjunc)on
and
transforma)on.
b)
Methods
of
internal
gene
duplica)on;
transposons,
gene
error
and
unequal
crossover.
a
Figure
1.
Proline
synthesis.
An
overview
of
proline
synthesis
and
the
involvement
of
proC
Figure
5.
Methods
overview.
The
en)re
process
involved
in
func)onal
complementa)on.
Figure
4.
Control
expected
results.
These
are
the
results
of
the
controls
and
the
proC
genes
given
they
are
adequate
for
viability.
Minimal
media
(MM),
MM
with
proline
and
MM
with
arabinose
plates
are
all
used
to
induce
different
effects.
Figure
7.
Func)onal
complementa)on.
a)
A.
vi7s
S4
results
for
func)onal
complementa)on
indicate
that
the
plasmid
with
proC-‐3168
was
sufficient
for
viability.
b)
A.
radiobacter
K84
results
for
func)onal
complementa)on
indicate
that
the
plasmid
with
arad
proC-‐12144
was
sufficient
for
viability.
Numbering
key
can
be
viewed
in
fig
8.
Figure
8.
Func)onal
complementa)on
and
analysis.
a)
Func)onal
complementa)on
with
all
plates
and
controls.
proC1-‐9900
showed
growth
in
minimal
media
with
arabinose
while
proC1-‐45470
showed
no
growth.
b)
Structural
comparison
of
proC1-‐
9900
(leh)
and
proC1-‐47450
(right)
to
see
if
structure
influences
viability
of
proC.
proC-‐4745
Figure
6.
New
vector
transforma)on
confirma)on.
Using
competent
cells
the
newly
created
vectors
for
proC1-‐65500,
proC1-‐39290
and
proC1-‐31690
two
from
A.
rhizogenes
A4
and
one
from
A.
vi)s
S4
respec)vely.
The
first
three
lanes
are
digest
from
proC-‐65500
followed
by
5
lanes
of
proC-‐39290
and
the
final
lane
of
proC-‐3168.
A
DNA
ladder
is
present
in
the
first
lane
to
compare
fragment
size.
The
top
band
in
the
digest
products
is
roughly
3.8
kb
and
the
products
for
the
inserts
are
all
roughly
around
.8
kb
in
size.
Figure
2.
Outcomes
of
gene
fixa)on.
There
are
three
main
possible
outcomes
aher
gene
fixa)on,
becoming
a
psuedogene,
subdividing
func)on
with
another
gene
or
gaining
a
gene
gene
func)on
en)rely.
b
a
b
a
b
proC1-‐9900
a