

Recommandé
Contenu connexe
Tendances
Tendances (20)
Similaire à Tejido nervioso
Similaire à Tejido nervioso (20)
Plus de Juan Opazo
Plus de Juan Opazo (20)
Tejido nervioso
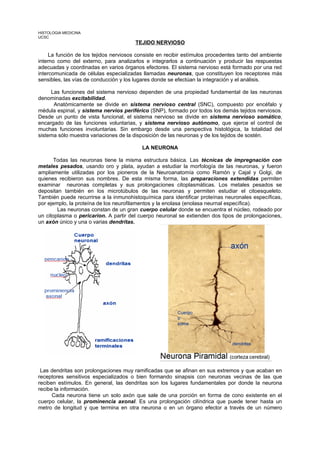
- 1. HISTOLOGIA MEDICINA UCSC TEJIDO NERVIOSO La función de los tejidos nerviosos consiste en recibir estímulos procedentes tanto del ambiente interno como del externo, para analizarlos e integrarlos a continuación y producir las respuestas adecuadas y coordinadas en varios órganos efectores. El sistema nervioso está formado por una red intercomunicada de células especializadas llamadas neuronas, que constituyen los receptores más sensibles, las vías de conducción y los lugares donde se efectúan la integración y el análisis. Las funciones del sistema nervioso dependen de una propiedad fundamental de las neuronas denominadas excitabilidad. Anatómicamente se divide en sistema nervioso central (SNC), compuesto por encéfalo y médula espinal, y sistema nervios periférico (SNP), formado por todos los demás tejidos nerviosos. Desde un punto de vista funcional, el sistema nervioso se divide en sistema nervioso somático, encargado de las funciones voluntarias, y sistema nervioso autónomo, que ejerce el control de muchas funciones involuntarias. Sin embargo desde una perspectiva histológica, la totalidad del sistema sólo muestra variaciones de la disposición de las neuronas y de los tejidos de sostén. LA NEURONA Todas las neuronas tiene la misma estructura básica. Las técnicas de impregnación con metales pesados, usando oro y plata, ayudan a estudiar la morfología de las neuronas, y fueron ampliamente utilizadas por los pioneros de la Neuroanatomía como Ramón y Cajal y Golgi, de quienes recibieron sus nombres. De esta misma forma, las preparaciones extendidas permiten examinar neuronas completas y sus prolongaciones citoplasmáticas. Los metales pesados se depositan también en los microtúbulos de las neuronas y permiten estudiar el citoesqueleto. También puede recurrirse a la inmunohistoquímica para identificar proteínas neuronales específicas, por ejemplo, la proteína de los neurofilamentos y la enolasa (enolasa neurnal específica). Las neuronas constan de un gran cuerpo celular donde se encuentra el núcleo, rodeado por un citoplasma o pericarion. A partir del cuerpo neuronal se extienden dos tipos de prolongaciones, un axón único y una o varias dendritas. Las dendritas son prolongaciones muy ramificadas que se afinan en sus extremos y que acaban en receptores sensitivos especializados o bien formando sinapsis con neuronas vecinas de las que reciben estímulos. En general, las dendritas son los lugares fundamentales por donde la neurona recibe la información. Cada neurona tiene un solo axón que sale de una porción en forma de cono existente en el cuerpo celular, la prominencia axonal. Es una prolongación cilíndrica que puede tener hasta un metro de longitud y que termina en otra neurona o en un órgano efector a través de un número
- 2. variable de pequeñas ramas con pequeñas dilataciones en sus extremos llamados botones terminales. Los potenciales de acción se originan en el cuerpo celular gracias a la integración de estímulos eferentes (que llegan); a continuación estos potenciales de acción viajan a lo largo del axón para influir en otras neuronas u órganos efectores. A menudo, se llama a los axones fibras nerviosas. En general, los cuerpos celulares de todas las neuronas se encuentran en el sistema nervioso central, salvo los de las neuronas sensitivas más primitivas y los de las neuronas efectoras terminales del sistema nervioso autónomo que, en ambos casos, se encuentran formando grupos llamados ganglios en localizaciones periféricas. Tipos básicos de neuronas En todo el sistema nervioso, las neuronas adoptan una gran variedad de formas que pueden agruparse en tres patrones principales, según la disposición del axón y las dendritas en relación con el cuerpo celular. Neurona multipolar, en la que numerosas dendritas se proyectan del cuerpo celular; las dendritas pueden salir todas de un mismo polo del cuerpo celular o nacer de todas las partes del mismo. En general, las neuronas intermedias, las de integración y las motoras siguen este patrón. Neuronas bipolares sólo tienen una dendrita, que sale del polo del cuerpo celular opuesto al origen del axón. Son escasas y actúan como receptores de los sentidos del olfato, la vista y el equilibrio. Neuronas seudounipolares, tienen una sola dendrita que nace junto al axón de un tallo común del cuerpo celular. Este tallo está formado por la fusión de la primera parte de la dendrita y del axón de una neurona de tipo bipolar, fusión que se produce durante el desarrollo embrionario. En general, los impulsos nerviosos viajan por las dendritas hacia el cuerpo neuronal (aferentes) mientras que por los axones viajan los impulsos que salen del cuerpo neuronal (eferentes). Las neuronas son células de diferenciación terminal y, en la práctica, cuando mueren no se regeneran. Se ha demostrado que las neuronas del encéfalo adulto pueden dividirse, pero el significado biológico se este hecho no se conoce. Sin embargo los axones y las dendritas sí pueden regenerar en caso de lesión, siempre que el cuerpo neuronal permanezca viable. Esta es la base de los injertos nerviosos usados para tratar las lesiones de los nervios periféricos. Ultraestructura de la neurona El núcleo es grande, redondeado u oval y habitualmente en el centro de la célula, rodeado por el pericarion. Como reflejo de la intensa actividad metabólica de la célula (y la consiguiente necesidad de sustituir a las proteínas alteradas), la cromatina está totalmente dispersa y el nucléolo es prominente. El citoplasma del cuerpo celular contiene grandes agregados de retículo endoplásmico rugoso (RER): la sustancia de Nissl. El RER se extiende por el interior de las dendritas pero no penetra en la prominencia axonal ni en el axón. Junto al núcleo se encuentra un aparato de Golgi difuso. El retículo endoplásmico liso no es una característica importante del pericarion, pero en las dendritas y el axón se encuentran túbulos, cisternas y vesículas claramente visibles. Las mitocondrias son numerosas en el pericarion, siendo más finas en el axón. Las neuronas tienen una gran actividad metabólica y consumen mucha energía para mantener el gradiente iónico a través de la membrana plasmática. Además, en el pericarion sintetizan sustancias neurotransmisoras o sus precursores, que son transportados a lo largo del axón hasta la sinapsis, donde son liberadas al recibir el estímulo adecuado.
- 3. Por todo el pericarion y a lo largo del axón y las dendritas existen numerosos filamentos intermedios (neurofilamentos) y microtúbulos, agrupados en haces paralelos. Los filamentos intermedios proporcionan sostén estructural y los microtúbulos intervienen en el transporte axonal de las sustancias neurotransmisoras, las enzimas y los componentes de las membranas y de otras partes de la célula. Fibras nerviosas mielinizadas y no mielinizadas En el sistema nerviosos periférico, todos axones están envueltos por células muy especializadas denominadas células de Schwann que les proporcionan un sostén tanto estructural como metabólico. En general, los axones de diámetros pequeños (es decir, los del sistema nervioso autónomo y las pequeñas fibras del dolor) están envueltas sólo por el citoplasma de las células de Schwann; se dice que estas fibras nerviosas son amielinicas. Las fibras de diámetro grande están rodeadas por un número variable de capas concéntricas de la membrana plasmática de la célula de Schwann que forman una vaina de mielina, por lo que se dice que estas fibras están mielinizadas. En el interior del sistema nervioso central, la mielinización es similar a la del sistema periférico, salvo por el hecho de que son las células llamadas oligodendrocitos las que forman las vainas de mielina. En todas las fibras nerviosas, la velocidad de conducción de los potenciales de acción es proporcional al diámetro de los axones; la mielinización aumenta mucho la velocidad de conducción del axón en comparación con la de las fibras no mielinizadas del mismo diámetro. Fibras nerviosas amielínicas: En la célula de Schwann se invaginan longitudinalmente uno o más axones, de forma que cada uno de ellos queda incluido en un canal revestido por la membrana plasmática y el citoplasma de la célula de Schwann. La membrana plasmática de ésta se adosa sobre sí misma a lo largo de la apertura del canal, produciendo un sellado eficaz del axón en un compartimiento extras celular rodeado por la célula de Schwann. La zona de aposición de la membrana plasmática de dicha célula recibe el nombre de mesaxón. Cada canal del interior de la célula de Schwann puede estar ocupado por más de un axón. Cada célula se extiende sólo durante un corto trayecto a lo largo del nervio y cuando termina, otra célula se hace cargo del revestimiento, mediante la íntima interdigitación de sus extremos. Fibra nerviosa mielinizada: En los nervios periféricos, la mielinización comienza con la invaginación de un sólo axón nervioso en una célula de Schwann, tras lo que se forma el mesaxón. A medida que la mielinización avanza, el mesaxón rota alrededor del axón, de forma que éste que cubierto por capas concéntricas del citoplasma y la membrana plasmática de la célula de Schwann A continuación el citoplasma desaparece, es desplazado y las capas internas de la membrana plasmática se fusionan entre sí, por lo que el axón acaba rodeado por múltiples capas de membrana que, en conjunto constituyen la vaina de mielina. Dada la alta concentración de lipoproteínas de la membrana plasmática, la mielina es una sustancia rica en lípidos. Al segmento de mielina producido por cada célula de Schwann se denomina internódulo y reviste al axón entre un nódulo de Ranvier. En el SNC, los responsables del proceso de mielinización son los oligodendrocitos, que siguen un proceso similar; sin embargo, un solo oligodendrocito forma múltiples internódulos de mielina, constituyendo a revestir un número de axones que puede llegar a 50. Nódulos de Ranvier e incisuras de Schmidt-Lanterman La vaina de mielina de un axón está formada por muchas células de Schwann (oligodendrocitos en el SNC), y cada célula de Schwann cubre sólo un segmento del axón. Entre las células de Schwann existen cortos intervalos en los que el axón no posee vaina de mielina; estas regiones reciben el nombre de nódulos de Ranvier. Sinapsis y uniones neuromusculares
- 4. Como en todas las células, las neuronas en reposo mantienen un gradiente iónico a través de la membrana plasmática, creando así un potencial eléctrico. La excitabilidad supone un cambio de la permeabilidad de la membrana como respuesta a los estímulos adecuados, de forma que el gradiente iónico se invierte y la membrana plasmática se despolariza; a continuación se produce una onda de despolarización llamada potencial de acción que se propaga por la membrana plasmática. A ello sigue un proceso de repolarización, por el que la membrana restablece rápidamente su potencial de reposo. En las sinapsis, los lugares de intercomunicación entre las neuronas adyacentes, la despolarización de una neurona induce la liberación de sustancias químicas transmisoras llamadas neurotransmisores, que indican un potencial de acción en la neurona adyacente. En el interior del sistema nervioso, las neuronas se disponen en forma de vías para la conducción de los potenciales de acción desde los órganos receptores a los efectores, a través de las neuronas de integración Los neurotransmisores no sólo intervienen en la transmisión de neurona a neurona, sino que también actúan como intermediarios químicos entre el sistema nervioso y los órganos efectores que poseen asimismo propiedades de excitabilidad. Los órganos efectores de las vías nerviosas voluntarias son, en general, los músculos esqueléticos mientras que los de las vías involuntarias suelen ser el músculo liso, el músculo cardiaco y las células mioepiteliales de algunas glándulas endocrinas. Las sinapsis son uniones intercelulares altamente especializadas que establecen comunicación entre las neuronas de cada vía nerviosa. Cada neurona establece sus comunicaciones a través de un número muy variable de sinapsis, dependiendo de su localización y de su función dentro del sistema nervioso. Las sinapsis pueden ser: sinapsis axodendríticas, sinapsis axomáticas e incluso sinapsis axoaxómicas. En cada sinapsis, la conducción de un impulso se hace en una sola dirección, pero la respuesta puede ser excitatoria o inhibitoria, dependiendo de la naturaleza funcional específica de las sinapsis y de su localización. El mecanismo de conducción del impulso nervioso implica la liberación de un neurotransmisor químico por una neurona, que difunde a través del estrecho espacio intercelular, para inducir la excitación o inhibición de otra neurona o célula efectora de la sinapsis. Los neurotransmisores ejercen sus efectos actuando sobre receptores específicos de la membrana plasmática de la célula opuesta. La naturaleza química de los neurotransmisores (NT) y la morfología de la sinapsis son muy variables en las distintas partes del sistema nervioso, pero los principios de la transmisión sináptica y la estructura básica de la sinapsis son similares. El axón termina en un ensanchamiento bulboso o botón terminal; separado de la membrana plasmática de la neurona o célula efectora opuesta por una hendidura intercelular, de 20-30, denominada hendidura sináptica. Los botones terminales no están mielinizados y contienen mitocondrias y vesículas que contienen neurotransmisores, las vesículas sinápticas de 50 nm. Aunque en el SNC existen muchos tipos de sustancias NT, en el sistema nervio periférico sólo se conocen dos tipos: acetilcolina y noradrenalina. Las vesículas sinápticas son transportadas desde el cuerpo celular, lugar de la síntesis de NT, hasta los botones sinápticos de la parte final del axón. Además, las vesículas también se forman en el botón sináptico por reciclado de las membranas. Las vesículas sinápticas se concentran en la membrana presinápticas y, cuando llega un potencial de acción entran el contacto con la membrana y liberan su contenido hacia la hendidura sináptica mediante exocitosis. El NT difunde a través de la hendidura sináptica y estimula a los receptores de la membrana postsináptica. En la sinapsis hay además varios mecanismos asociados como enzimas hidrolíticas y oxidativas que, entre los impulsos nerviosos sucesivos, inactiva al neurotransmisor liberado. El transmisor también puede volver al botón terminal, donde es reciclado en nuevas vesículas sinápticas. El citoplasma situado bajo la membrana postsináptica suele contener una banda de fibras finas, la red postsináptica, que puede a estructuras de tipo desmosoma para mantener la integridad de la sinapsis. Esta densidad
- 5. postsináptica contribuye a la estabilidad estructural de las membranas pre y post sinápticas íntimamente adosadas. Placas motoras terminales Existen uniones intercelulares que conectan a las neuronas con sus células efectoras, como fibras musculares, y en este caso las sinapsis se llaman uniones neuromusculares o placas motoras terminales. La estructura básica de las placas motoras terminales del músculo esquelético es igual a las de las demás sinapsis, pero con algunas características especiales. Una neurona motora puede inervar desde unas pocas a más de mil fibras musculares, dependiendo de la precisión del movimiento del músculo; la neurona motora constituye, junto con las fibras musculares que inerva, la unidad motora. La placa motora Terminal ocupa una depresión en la superficie de la célula muscular, descrita como placa plantar, cubierta por una extensión del citoplasma de al menos una de las células de Schwann que rodean al axón. La membrana postsináptica de la unión neuromuscular desarrolla unos pliegues profundos que se forman hendiduras sinápticas secundarias paralelas. La membrana presináptica situada por encima también es irregular y el citoplasma contiene numerosas vesículas sinápticas. En la placa plantar de la fibra muscular se produce una concentración de mitocondrias y de núcleos de la célula muscular. El neurotransmisor de las uniones neuromusculares somáticas es la acetilcolina, cuyos receptores se concentran en los márgenes de las hendiduras sinápticas secundarias. La enzima hidrolítica acetilcolinesterasa ocupa la parte más profunda de las hendiduras que se asocian a la lámina externa e intervienen en la desactivación del neurotransmisor entre los impulsos nerviosos sucesivos. TEJIDOS NERVIOSOS PERIFÉRICOS Los tejidos nerviosos periféricos son estructuras anatómicas que pueden estar formadas por una combinación de fibras nerviosas aferentes y eferentes o por fibras pertenecientes a los sistemas nerviosos somático o autónomo. Los cuerpos celulares de las fibras que viajan en los nervios periféricos se encuentran en el SNC o en los ganglios periféricos. Cada nervio periférico está formado por uno o más haces de fibras nerviosas. En el interior de los fascículos, cada fibra nervosa individual, revestida por la célula de Schwann, está rodeada por un delicado tejido de sostén laxo y vascular denominado endoneuro. A su vez cada fascículo estaá rodeado por una capa condensada de tejido de colágeno denominado perineuro. En los nervios periféricos formados por más de un fascículo, existe otra capa de tejido colágeno laxo llamado epineuro que mantiene unidos los fascículos y que se condensa en la periferia para formar una fuerte vaina cilíndrica. Los nervios periféricos reciben una rica irrigación sanguínea a través de numerosos vasos penetrantes que proceden de los tejidos adyacentes y de las arterias que acompañan a los nervios. Los vasos de mayor calibre siguen un trayecto longitudinal en el compartimiento rodeado por el perineuro y el epineuro y forman una abundante red capilar en el endoneuro. Un gran número de anastomosis garantiza la irrigación adecuada. Corte longitudinal Ganglio raquídeo
- 6. Son pequeños agregados bien definidos de cuerpos celulares neuronales, localizados fuera del SNC. Se encuentran en las raíces nerviosas posteriores de la médula espinal, en la zona por la que atraviesan los agujeros intervertebrales. Contienen los cuerpos celulares de neuronas sensitivas primarias, de forma seudo-unipolar. El ganglio está rodeado por una cápsula de tejido de sostén condensado que se continúa con las vainas perineural y epineural del nervio periférico asociado. Ganglio simpático Los ganglios simpáticos tienen una estructura similar a la de los ganglios sensitivos somáticos. Las neuronas son multipolares, por lo que se encuentran más esparcidas, separadas por numerosos axones y dendritas, muchas de las cuales pasan por el ganglio sin establecer sinapsis. Ganglio parasimpático Los cuerpos celulares de las neuronas efectoras terminales del sistema nervioso parasimpático se encuentran en el interior del propio órgano efector o en su vecindad. Pueden formar ganglios bien organizados de tamaño medio (como el ganglio óptico), pero lo más frecuente es que se agrupen en pequeños ganglios dispersos por el tejido de sostén. Como todas las neuronas, las células ganglionares se reconocen por su núcleo y gran tamaño, con cromatina dispersa y nucléolo prominente, y por su amplio citoplasma basófilo. También en estos ganglios las neuronas están rodeadas por numerosas células de sostén pequeñas y por fibras nerviosas aferentes y eferentes. TEJIDOS NERVIOSOS CENTRALES El SNC está formado por el encéfalo y la médula espinal; cada una de estas partes puede dividirse microscópicamente en áreas de sustancia gris y de sustancia blanca. En la sustancia gris se encuentran casi todos los cuerpos neuronales y sus fibras asociadas axones. La sustancia blanca está formada por haces de fibras nerviosas en los que un número importante de axones están mielinizados, siendo la mielina la que confiere la apariencia blanca al tejido fresco. En tejido nervioso central consta de un enorme número de neuronas y sus prolongaciones incluidas en una masa de células de sostén que, en conjunto reciben el nombre de neuroglia, que abarca a todas las células del SNC que no son neuronas. El tejido nervioso central propiamente dicho no tiene tejido de sostén colágeno, que se limita a la inmediata vecindad de los vasos sanguíneos penetrantes y a las meninges que recubren la superficie externa. La neuroglia, que forma alrededor de la mitad de la masa total del SNC, está formada por células muy ramificadas que ocupan los espacios existentes entre las neuronas; el material extracelular es muy escaso en el SNC. Entre la neuroglia y las neuronas existen unas íntimas relaciones funcionales de forma que aquella proporciona el sostén mecánico y metabólico que necesitan las segundas. Se reconocen cuatro tipos principales de neuroglia: - Oligodendrocitos, son el equivalente a las células de Schwann, son responsables de la elaboración de las vainas de mielina. - Astrocitos, son células muy ramificadas que llenan los intersticios que quedan entre las neuronas, sus prolongaciones y los oligodendrocitos. Proporcionan un sostén mecánico e intervienen en el intercambio de metabolitos entre las neuronas y el sistema vascular. Además forman parte de la barrera hematoencefática. Los astrositos también desempeñan un papel importante en la reparación del SNC tras una lesión o daño por enfermedad. - Microglia, es la representante en el SNC del sistema monolito-macrófago y sus funciones son efectivas e inmunológicas.
- 7. - Células ependimarias, forman un epitelio especializado que reviste los ventrículos y el canal medular. Sustancia gris Aunque el tamaño y la morfología de las neuronas son muy variables de unas regiones a otras del encéfalo, suelen ser reconocibles por su gran núcleo o nucléolo prominentes y cromatina dispersa y por un amplio citoplasma basófilo granular, del pueden verse salir una o varias prolongaciones. En el SNC maduro, los oligodendrocitos tienen núcleos pequeños, rodeados y condensados en su citoplasma. Los oligondendrocitos de la sustancia gris no sólo se encuentran dispersos entre los cuerpos de las células nerviosas junto con los astrocitos, sino que tienden también a agregarse en torno a dichos cuerpos neuronales. Los núcleos tanto de las neuronas como de la neuroglia están rodeados por una capa de axones y dendritas que salen y convergen en las neuronas. Este tejido recibe el nombre de neuropilo. La mayor parte de las fibras del neuropilo carecen de mielina (están demasiado cerca de los cuerpos celulares neuronales), lo que justifica su eosinofilia. Astrocitos Estas células las más numerosas de todas las gliales de la sustancia blanca, tienen largas prolongaciones ramificadas que ocupan la mayor parte de los espacios interneuronales. En la sustancia gris, muchas de las prolongaciones astrocitarias terminan en expansiones adyacentes a las regiones no sinápticas de las neuronas. Otros procesos de los mismos astrocitos terminan sobre las membranas basales de los capilares de forma que estos pies perivasculares cubren gran parte de la superficie de las membranas basales capilares. Prolongaciones similares revisten la membrana basal situada entre el SNC y la capa más interna de las meninges, la piamadre formando una barrera relativamente impermeable llamada glia limitante. De esta forma, los astrocitos de la sustancia gris intervienen en el intercambio metabólico entre las neuronas y la sangre y regulan la composición del medio intercelular del SNC. Todos los astrocitos contienen haces de filamentos intercelulares y microtúbulos, especialmente prominentes en los astrocitos de la sustancia blanca, que contienen menos prolongaciones citoplasmáticas rectas y a los que se conoce como astrocitos fibrosos. Por el contrario, los de la sustancia gris poseen numerosas prolongaciones citoplasmáticas cortas y muy ramificadas; son los astrocitos protoplasmáticos. Los filamentos intermedios consisten en una proteína característica de los astrocitos denominado proteína ácida fibrilar gliar (PAFG). Oligodendrocitos El nombre de oligodendrocito se debe a los primeros neurohistólogos, que utilizaban métodos de impregnación con metales pesados que mostraban a estas células con un pequeño número de prolongaciones cortas y ramificadas (griego: oligos = poco: dendron = árbol). Hoy se sabe que son las responsables de la mielinización de los axones en el SNC y que las prolongaciones que se describían corresponden a cortos pedículos que conectan el cuerpo celular con las vainas de mielina. De hecho un solo oligodendrocito puede contribuir a la mielinización de hasta 50 axones, pertenecientes al mismo o a distintas haces de fibras. A la inversa, cualquier axón necesita los servicios de numerosos oligodendrocitos distintos, ya que los espacios internodales existentes a lo largo de su trayecto dependen de células distintas. El mecanismo de la formación de la vaina de mielina es muy similar al de las células de Schwann en el nervio periférico. Por tanto los oligodendrocitos son el tipo de neuroglia predominante en la sustancia blanca, además de ser también muy abundantes en la sustancia gris. Por otra parte, estas células se agrupan estrechamente alrededor de los cuerpos de las neuronas en la sustancia gris, donde parecen ejercer una función de sostén similar a la de las células satélite que rodean a los cuerpos neuronales en los ganglios periféricos. La formación de la vaina de mielina se inicia en el SNC del embrión humano hacia los 4 meses de edad gestacional y cuando el niño tiene un año de vida la mayor parte de las vainas han comenzado ya, al menos, a formarse. A partir de ese momento, continúan depositándose capas sucesivas, de forma que la vaina alcanza su grosor definitivo cuando el individuo llega a la madurez física. Se describen tres tipos de oligodendrocitos, claros (con capacidad de división y son muy activos en la formación de vainas de mielina, por lo que predominan en el feto y el recién nacido), medios (representan la forma inmadura que interviene en el crecimiento y maduración de la vaina de mielina)y oscuros (son la forma predominante en el SNC maduro). En el SNC maduro se encuentran algunas formas claras y medias, revelando la existencia de un cierto recambio lento y constante, con capacidad para remielinizar en caso necesario (ej. enfermedades desmielinizantes como la esclerosis múltiple).
- 8. Microglia Formada por células pequeñas de origen mesenquimal que penetran en el SNC en una etapa tardía del desarrollo fetal. Tienen núcleos alargados e irregulares y citoplasma relativamente escaso, que forman finas prolongaciones muy ramificadas. Son difíciles de identificar en preparaciones corrientes. En respuesta a lesiones, las células de la microglia se transforman en grandes elementos fagocitarios ameboides, por lo que se las considera representantes del sistema de defensa. En circunstancias normales el tráfico de linfocitos es muy escaso en el SNC, pero aumenta mucho en los procesos inflamatorios. Sin embargo, el número de macrófagos es considerable en el espacio que rodea a los capilares, aunque permanecen separados del SNC propiamente dicho por los pies perivasculares de los astrocitos. Epéndimo Las células ependimarias forman el revestimiento epitelial simple de los ventrículos y del canal medular. Estas células, de forma cúbica o cilíndrica baja, se encuentran íntimamente unidas por sus superficies luminares mediante complejos de unión. A diferencia de otros epitelios las células ependimarias no descansan sobre una membrana basal, sino que sus bases se afinan y después se dividen en finas ramas que penetran en la capa subyacente de prolongaciones derivadas de los astrocitos. En la superficie luminal poseen un número variable de cilios que podrían intervenir en la propulsión del líquido cefalorraquídeo en el interior de los ventrículos. Plexo coroideo Es una estructura vascular que surge de la pared de cada uno de los cuatro ventrículos del encéfalo y es la responsable de la producción del líquido cefalorraquídeo (LCR). Este drena en las cavidades ventriculares comunicantes a través de tres canales, que conectan los cuatro ventrículos con el espacio subaracnoideo situado en torno al SNC. El LCR se produce a un ritmo constante y se reabsorbe desde el espacio subaracnoideo al seno venoso sagital superior a través de proyecciones digitiformes, denominadas vellosidades aracnoideas. Gracias a él, el SNC está suspendido en un medio líquido que circula constantemente y que actúa absorbiendo los golpes. Cada plexo coroideo es una masa de capilares que se proyectan hacia el ventrículo y que están revestidos por células ependimarias modificadas. Desde las superficies luminales de las células epiteliales coroideas se proyectan grandes microvellosidades bulbosas y sus citoplasmas contienen numerosas mitocondrias, lo que indica que la elaboración del LCR es un proceso activo. Los capilares de los plexos coroideos son grandes, de pared fina y a veces fenestrados. El modo de secreción del LCR supone la secreción activa de iones de sodio por las células del epitelio coroideo hacia el LCR, seguida de un movimiento pasivo de agua desde los capilares. Las uniones íntimas continuas (zonula occludens) forman una barrera hematoencefálica que impide el paso de casi todas las demás moléculas. La glucosa (cuya concentración es aproximadamente 70% de las plasmática) y pequeñas cantidades de proteínas son componentes normales del LCR, pero se desconocen como llegan a él. MENINGES El encéfalo y la médula espinal están revestidos por tres capas de tejido de sostén que, en conjunto, reciben el nombre de meninges. Piamadre: cubre el tejido nervioso y contiene fibras de colágeno, finas fibras elásticas y fibroblastos, separados de las prolongaciones de los astrocitos subyacentes por una membrana basal. Esta última está completamente cubierta por las prolongaciones de los astrocitos y las dos capas forman la glia limitante impermeable. Aracnoides: se encuentra sobre la piamadre, es una capa de tejido fibroso más gruesa. Debido a la continuidad estructural entre ambas, a menudo se considera que forman una unidad denominada pia-aracnoides o leptomeninge. Espacio subaracnoideo: es el espacio entre la pia y la aracnoides y en determinados lugares forman grandes cisternas. El espacio subaracnoideo comunica con el sistema ventricular por tres agujeros y el LCR circula continuamente desde los ventrículos a dicho espacio. Las superficies opuestas de la pia y la aracnoides y las fibras que se comunican están revestidas por células mesoteliales aplanadas. La superficie externa de la aracnoides también está revestida por mesotelio. Las arterias y las venas que pasan desde y hacia el SNC atraviesan el espacio subaracnoideo laxamente unidas a la pia y están revestidas por el mesotelio subaracnoideo. Los grandes vasos que se introducen en el tejido nervioso están rodeados por una delicada envoltura de piamadre. Entre los vasos penetrantes y la pia existe un espacio perivascular continuo con el espacio subaracnoideo en algunos animales pero no en el hombre, en el que el epitelio de la pia se mezcla con la adventicia de los vasos cuando ésta penetra en el encéfalo, separando los espacios perivascular y subaracnoideo.
- 9. Duramadre: por fuera de la aracnoides, es una densa capa fibroelástica, revestida en su superficie interna por mesotelio. La dura está íntimamente adosada pero no fusionada a la aracnoides, de forma que entre ambas puede desarrollarse un espacio potencial, el espacio subdural. En el encéfalo, la duramadre se une al periostio del cráneo, mientras que en la médula espinal está suspendida del periostio del canal raquídeo por los ligamentos dentriculados, existiendo por tanto un espacio epidural ocupado por tejido fibroadiposo laxo y plexos venosos. Los estudios de perfusión demuestran que los capilares del SNC son impermeables para determinados componentes del plasma, sobre todo para las grandes moléculas, de modo que forman una barrera hematoencefálica. El endotelio capilar desempeña un papel esencial, ya que las uniones entre las células endoteliales están selladas; las células endoteliales no muestran pinocitosis o ésta es muy escasa. Las membranas de la superficie luminal contiene varias enzimas que destruyen los metabolitos neurotóxicos y las sustancias humorales neuroactivas. El mantenimiento de la barrera endotelial parece estar controlado por los pies astrocitarios. La barrera hematoencefálica proporciona a las neuronas un medio bioquímico y metabólico relativamente constante, las protege frente a las toxinas endógenas y exógenas y los agentes infecciosos y las aísla de los neurotransmisores circulantes y de otros agentes humorales. Sin embargo, los capilares del plexo coroideo, la hipófisis y la glándula pineal, así como el centro del vómito del hipotálamo, no tienen esta barrera, como conviene para sus variadas funciones. RECEPTORES SENSORIALES Los receptores sensoriales son terminaciones nerviosas o células especializadas que convierten (traducen) los estímulos procedentes de los ambientes externo o interno en impulsos nerviosos aferentes; estos impulsos llegan al SNC donde ponen el marcha las respuestas voluntarias o involuntarias adecuadas. Una clasificación funcional ampliamente utilizada los divide en tres grupos: - extoreceptores, son los que responden a estímulos procedentes del exterior del organismo e incluyen a los receptores del tacto, la presión superficial, la presión profunda, el dolor cutáneo, la temperatura, el olor, el sabor, la vista y el oído. - Propioceptores, se encuentran en el sistema esquelético y proporcionan información consciente e inconsciente de la orientación, la posición del esqueleto, la tensión y el movimiento; estos receptores son el aparato vestibular del oído, los órganos tendinosos y los huesos neuromusculares. - Interoreceptores, responden a estímulos procedentes de las vísceras y son los quimiorreceptores de la sangre, los barorreceptores vasculares, los receptores del estado de distensión de las vísceras huecas y receptores para sensaciones tan nebulosas como el dolor visceral, el hambre, la sed, el bienestar y el malestar. La estructura de los receptores que intervienen en algunas de las modalidades sensitivas es mal conocida. Desde un punto de vista morfológico, pueden clasificarse en: simples (meramente terminaciones nerviosas libres, ramificadas o no ramificadas, como las responsables del dolor cutáneo y la temperatura) y compuestos (existe una organización con los tejidos no nerviosos
- 10. asociados que complementan la función del receptor neural propiamente dicho; el grado de organización puede oscilar entre una mera encapsulación a una estructura muy compleja como sucede en el ojo o el oído). Terminaciones nerviosas libres Las terminaciones nerviosas libres son las formas más sencillas de receptores sensitivos y consisten sólo en numerosas ramas terminales pequeñas de fibras nerviosas aferentes. Se encuentran en los tejidos de sostén de todo el organismo, donde contribuyen a distintas modalidades sensoriales relativamente simples, como la temperatura, el tacto y el dolor. Las fibras aferentes tiene diámetros relativamente pequeños, con velocidades de conducción lentas; aunque algunas de estas fibras están mielinizadas, las terminaciones no tienen mielina. En la piel, aparecen a lo largo de la unión dermo-epidérmica. Algunas íntimamente asociada a células no neuronales llamadas células de Merkel, que se encuentran diseminadas por la capa de la epidermis. El citoplasma de la célula de Merkel adyacente contiene vesículas con características ultraestructurales similares a las de las sinapsis (pero por el momento no se ha demostrado la presencia de neurotransmisores), que están conectadas a fibras mielínicas de gran diámetro y serían las responsables del tacto. En los folículos de los pelos finos y gruesos existen diversas organizaciones de terminaciones nerviosas libres que actúan como receptores táctiles. Corpúsculos de Meissner Los corpúsculos de Meissner son pequeños receptores sensoriales encapsulados que se encuentran en la dermis cutánea, especialmente en los pulpejos de los dedos de las manos, las plantas de los pies, los pezones, los párpados, los labios y los genitales. Intervienen en la recepción del tacto fino discriminativo. Corpúsculos de Pacini Son grandes receptores sensitivos encapsulados que responden a la presión y al tacto grosero, a la vibración y a la tensión y que se encuentran en las capas más profundas de la piel, los ligamentos y las cápsulas articulares, en algunas membranas serosas, el mesenterio, algunas vísceras y en algunas zonas erógenas. Miden entre 1 y 4 mm y al corte presentan un aspecto parecido a una cebolla. Están formados por una delicada cápsula que rodea a muchas laminillas concéntricas de células aplanadas (probablemente células de Schwann modificadas), separadas por espacios de líquido intersticial y delicadas fibras de colágeno. El núcleo está formado por una única fibra nerviosa grande, no ramificada ni mielinizada, con varias terminaciones ensanchadas que se mielinizan cuando salen del corpúsculo. La distorsión de los corpúsculos de Pacini produce un estímulo mecánico amplificado en el centro que se traduce en un potencial de acción en la neurona sensitiva. Se describen otros dos mecanorreceptores encapsulados simples, los corpúsculos de Ruffini que son robustas estructuras fusiformes más frecuentes en las plantas de los pies, y los bulbos terminales de Krause, receptores delicados que se encuentran en el revestimiento de la orofaringe y en la conjuntiva ocular. Huso neuromuscular Son receptores de distensión que se encuentran en los músculos esqueléticos y son los responsables de la regulación del tono muscular a través del reflejo de distensión medular. Son más numerosos en los músculos que intervienen en los movimientos finos de precisión. Se estimulan por el estiramiento de las fibras intrafusales que tiene lugar cuando la masa muscular extrafusal se distiende. Este estímulo despierta una contracción refleja de las fibras musculares a través de las neuronas motoras largas (alfa) de un arco reflejo medular simple de dos neuronas. La contracción de la masa muscular contrarresta el estímulo de distensión de los receptores y el equilibrio se restablece. La sensibilidad del huso neuromuscular está modulada por los centros superiores a través de las neuronas motoras pequeñas (gamma) del sistema extrapiramidal. Estas neuronas motoras gamma inervan las porciones estriadas de las fibras intrafusales controlando su estado de contracción. La contracción de las fibras intrafusales aumenta la sensibilidad de los receptores intrafusales a la distensión de la masa extrafusal.