Motor system overview
•Télécharger en tant que PPTX, PDF•
3 j'aime•6,093 vues

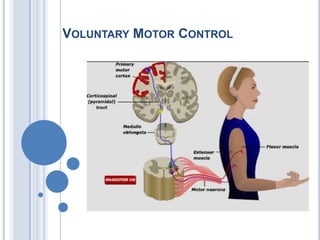
Voluntary motor control involves multiple brain areas that work together. The primary motor cortex directly controls spinal motor neurons and codes for movement force and direction. Other areas like the dorsal premotor cortex encode sensorimotor transformations for visual and sensory cues. The basal ganglia and cerebellum help modulate and coordinate movement through disinhibition of pathways. Feedback from muscles and joints via sensory neurons also helps regulate movement.

Recommandé
Contenu connexe
Tendances
Tendances (20)
Similaire à Motor system overview
Similaire à Motor system overview (20)
Plus de PS Deb
Plus de PS Deb (20)
Dernier
Dernier (20)
Motor system overview
- 2. Brain vs. Heart Hippocrates 460-377 BC, “Men ought to know that from the brain and from the brain only arise our pleasures, joys, laughter and jests, as well as our sorrows, pains, grieves and tears”. Aristotle 384-322 B.C “the heart as the organ of thinking, of perception and feelings,” “brain could cool the passion of heart” 387 B.C. - Plato teaches at Athens. Believes brain is seat of mental process
- 3. Brain as hollow organ: 335-280 BC Herophilus Ventricles are seat of human intelligence
- 4. Galen 130-200 AD 177 - Galen lecture On the Brain
- 5. Leonardo Da Vinci April 15, 1452 – May 2, 1519
- 6. Andreas Vesalius (1514-1564 CE)
- 7. Descartes, Brain and Mind 1649 - Rene Descartes describes pineal as control center of body and mind
- 8. 1664 - Thomas Willis 1664 - Publishes Cerebrianatome (in Latin) 1681 - coins the term Neurology
- 9. Bioelectricity 1791 - Luigi Galvani publishes work on electrical stimulation of frog nerves 1809 - Luigi Rolando uses galvanic current to stimulate cortex
- 10. Phrenology Franz Joseph Gall 1806 The theory of Gall and Spurzheim is ... an instance of a theory which, while essentially wrong, was just enough right to further scientific thought…Edwin Boring
- 11. 1824 - Marie-Jean-Pierre Flourens
- 12. 1825 - Jean-BaptisteBouillaud Presents cases of loss of speech after frontal lesions
- 13. Pierre Gratiolet
- 14. Phinease Gage (1848) On 13th Sept 1848 a railroad worker hard working, diligent, reliable, responsible, intelligent, good humored, polite god fearing, family oriented foreman Following an explosion iron bar drove into frontal lobe 1. He becomes unreliable and fails to come to work and when present he is "lazy." 2. He has no interest in going to church, constantly drinks alcohol, gambles, and "whores about." 3. He is accused of sexually molesting young children. 4. He ignores his wife and children and fails to meet his financial and family obligations. 5. He has lost his sense of humour. 6. He curses constantly and does so in inappropriate circumstances. 7. Died of status epilepticus in 1861
- 15. Lobar Localization Paul Broca 1868 1861 - discusses cortical localization
- 16. 1870, Hitzig and Fritsch Electrically stimulated various parts of a dog's motor cortex. They observed that depending on what part of the cortex they stimulated, a different part of the body contracted. Then they found that if they destroyed this same small area of the cortex, the corresponding part of the body became paralyzed. This is how it was discovered that every part of the body has a particular region of the primary motor cortex that controls its movement.
- 17. 1870 - David Ferrier
- 18. John Hughlings Jackson Described seizure as cerebral event not medulla on clinicopathological observation of head injury patients.
- 19. Discovery of Neuron Ramony Cajal and Camillo Golgi 1906 Noble
- 21. 1906 - Sir Charles Scott Sherrington 1906 - Sir Charles Scott Sherrington publishes The Integrative Action of the Nervous system that describes the synapse and motor cortex
- 22. 1957 - W. Penfield
- 23. Control of voluntary movement Execution Planning Basal Ganglia Association cortex Premotor + Motor cortex Movement Idea Lateral cerebellum Intermediate Cerebellum
- 24. Organization of Motor system
- 25. Organization of Motor Nervous System
- 27. Functional Organization of the Primary Motor Cortex
- 29. The major inputs to the motor cortex in monkeys
- 32. Convergence of Motor Control on the Anterior Motor Neuron
- 33. Experimental apparatus developed to record the activity of single neurons in awake primates trained to perform specific movements : Ed Evarts 1960
- 34. Direct corticospinal control of motor neurons is necessary for fine control of the digits
- 35. Motor Cortical Cell Firing with Force Generated
- 36. Corticomotoneuronal (CM) cell is active depends on the motor task
- 37. Activity in Individual Neurons of the Primary Motor Cortex Is Related to Muscle Force and Direction of Movement
- 38. Spike Triggered Averaging 1970
- 39. Directional tuning of an upper motor neuron in the primary motor cortex
- 40. Different areas of cortex are activated during simple, complex, and imagined sequences of finger movements (Xenon PET)
- 41. Cell activity in the motor cortex depends on whether a sequence of movements is guided by visual cues or by prior training
- 42. A set-related neuron in the dorsal premotor area becomes active while the monkey prepares to make a movement to the left
- 43. The visuomotor transformations required for reaching and grasping involve two different pathways
- 44. Individual neurons in the ventral premotor area fire during specific hand actions only
- 45. Mirror Neurons A. Activity in the neuron as the monkey observes another monkey make a precision group.B. Activity in the same neuron as the monkey observes the human experimenter make the precision grip.C. Activity in the same neuron as the monkey itself performs a precision grip. (From Rizzolotti et al 1996.)
- 46. Different areas of cortex are activated during simple, complex, and imagined sequences of finger movements (Xenon PET)
- 47. The Somatotopic Organization of the Motor Cortex Is Plastic
- 48. As a movement becomes more practiced, it is represented more extensively in primary motor cortex
- 49. Summary Primary Motor Cortex: Codes force and direction of movement Spinal motor neuron are directly under control for precise movement. Dorsal Premotor Cortex Movement related neuron encodes sensorimotor transformation for visual and sensory cue Fire before movement Ventral Premotor Cortex Encodes learned motor act fire before movement All cortical neurons are adaptable and plastic
- 51. Modulation of Movement by the Basal Ganglia
- 52. Motor components of the human basal ganglia
- 53. Anatomical Organization of Basal Ganglia Input
- 54. Output of Basal Ganglia
- 55. Disinhibition in the direct and indirect pathways through the basal ganglia
- 57. Input of Cerebellum
- 60. Neocerebellum
- 61. The spinocerebellum contains two somatotopic neural maps of the body
- 62. Motor modulation by the cerebrocerebellum
- 65. Spinal Cord: Grey matter
- 66. Motor Unit
- 69. Mechanism of Muscle contraction
- 70. Muscle Spindle
- 71. Stretch Reflex
- 75. Feedback Inhibition: Golgi Tendon Organ
- 76. Alpha and gamma motor neurons are coactivated during voluntary movements
- 77. Spinal Animal
- 78. Sensory Feedback for walking
- 79. Locomotor Center in Cat
- 80. Thanks
Notes de l'éditeur
- In the late 1820s Gall's ideas were subjected to experimental analysis by the French physiologist Pierre Flourens. By systematically removing Gall's functional centers from the brains of experimental animals, Flourens attempted to isolate the contributions of each “cerebral organ” to behavior. From these experiments he concluded that specific brain regions were not responsible for specific behaviors, but that all brain regions, especially the cerebral hemispheres of the forebrain, participated in every mental operation. Any part of the cerebral hemisphere, he proposed, was able to perform all the functions of the hemisphere. Injury to a specific area of the cerebral hemisphere would therefore affect all higher functions equally.In 1823 Flourens wrote: “All perceptions, all volitions occupy the same seat in these cerebral) organs; the faculty of perceiving, of conceiving, of willing merely constitutes therefore a faculty which is essentially one.” The rapid acceptance of this belief (later called the aggregate-field view of the brain) was based only partly on Flourens's experimental work. It also represented a cultural reaction against the reductionist view that the human mind has a biological basis, the notion that there was no soul, that all mental processes could be reduced to actions within different regions in the brain!Eur Neurol. 2009;61(5):311-4. Epub 2009 Mar 17.Marie-Jean-Pierre Flourens (1794-1867) and cortical localization.Pearce JM.Department of Neurology, Hull Royal Infirmary, Hull, UK.AbstractThe child prodigy Marie-Jean-Pierre Flourens received his medical degree at Montpellier when aged 19. As a young promising physician Flourens was asked to investigate Gall's controversial views on cerebral localization. To test Gall's assertions, Flourens developed ablation as a procedure to explore the workings of the brain. By removing anatomically defined areas of the brain of an animal and watching its behaviour, he thought he might localize certain functions. Flourens did not favour the idea of cerebral localization and concluded that the brain functioned as a whole and thus arose the concept of 'cerebral equipotentiality'. This culminated in his 1824 Recherchesexpérimentalessur les propriétés et les fonctions du systèmenerveux. His techniques were, however, crude and imperfect, and his experiments were mainly on birds. Much criticism and debate ensued. A gifted man, Flourens also advanced the physiology of the vestibular apparatus and described the anaesthetic properties of ether.IntroductionBorn on 15 April 1794, in Maureilhan, near Béziers, France, Marie-Jean-Pierre Flourens (fig. 1) was something of a child prodigy. He enrolled at the famed Faculté de Médecine at Montpellier when he was only 15 years old and received his medical degree aged 19. Shortly thereafter, while Franz Gall (1758–1828) was at the height of his career in Paris, Flourens himself moved to the capital and under Georges Cuvier started to investigate brain functions and localization [1]. Gall’s work was questioned in some circles, and the Paris Academy of Sciences, acting on order of the Emperor Bonaparte, asked Flourens to investigate the matter. On the basis of his 1824 Recherchesexpérimentalessur les propriétés et les fonctions du systèmenerveux [2], he was elected to membership and eventually on Cuvier’s dying request was appointed Perpetual Secretary of the Académie des Sciences, to become one of France’s most revered scientists (fig. 2).Before Flourens began his animal experiments, Gall had developed relatively new ideas of brain function [3], which proved important:‘that the cerebral cortex represented the highest level of the brain and that the development of this area characterised mammals and man.’He tried to analyze and localize cerebral functions physiologically. His work moved ‘away from the concept of the insubstantial Cartesian soul to the concept of more material nerve function’, and in doing so ‘was wrong only in detail and in respect of the enthusiasm of its supporters’ [4]. His later ideas of phrenology [5] brought much of his work into disrepute, and physicians neglected his earlier significant advances. Nevertheless, as Edwin Boring wrote: ‘The theory of Gall and Spurzheim is ... an instance of a theory which, while essentially wrong, was just enough right to further scientific thought’ [6]. To test Gall’s assertions, Flourens developed ablation as a procedure to explore the workings of the brain. By removing anatomically defined areas of the brain of an animal and watching its behaviour, he thought he might localize certain functions. For example, by removal of the cerebellum, the animal could no longer coordinate movements. In 1823, he stated that the cerebellum regulates motor activity and a year later provided details of his ablation experiments on birds and lower vertebrates to study behaviour. By making discrete lesions in the medulla, near the posterior ventricle, he reported (cited in Changeux, p. 16, [7]):‘he disturbed certain vital functions, such as respiration.’Flourens did not favour the idea of cerebral localization and concluded that the brain functioned as a whole when he wrote:‘a large section of the cerebral lobes can be removed without loss of function. As more is removed, all functions weaken and gradually disappear. Thus the cerebral lobes operate in unison for the full exercise of their functions ... The cerebral cortex functioned as an indivisible whole ... [housing] an ‘‘essentially single faculty’’ of perception, judgement and will ... the last refuge of the soul’ (Flourens, cited by Changeux, p. 17, [7]).However, Flourens’s experimental technique was not accurate, and his behavioural studies were both crude and rudimentary for the adequate investigation of the faculties recently catalogued by Gall and Johann Spurzheim (1776–1832). Flourens related loss of function to the extent of damage. However, he adopted a holistic concept, with an understanding that the brain was composed of areas of specific functions, but specific areas interacted to work as a whole:‘All sensations, all perceptions, and all volition occupy concurrently the same seat in these organs. The faculty of sensation, perception, and volition is then essentially one faculty’ [2].Thus arose the concept of ‘cerebral equipotentiality’.While previous researchers had lesioned the brain through a trephine that made it impossible to localize damage accurately or to track haemorrhage, Flourens completely uncovered and isolated that portion of the brain to be removed. He tried to minimize operative trauma and post-operative bleeding and infection which might confound analysis. His ablation experiments led him to conclude the presence of a motor centre in the medulla oblongata and coordination in the cerebellum. Although his treatment of sensation was still rather confused, by the time the second edition of the Recherchesexpérimentales (1842) appeared, Flourens had clearly distinguished sensation and perception, treating perception as the appreciation of the meaning of a sensation. He thought that several sub-cortical structures were responsible for sensory function. However, his slicing of the hemispheres produced diffuse damage with resulting impairments of varied degree involving perception, intellect and will. The resulting damage varied with the extent of the lesion. After small lesions function might be restored, but total ablation caused permanent loss of function. Flourens concluded that while sensory-motor functions were localized to sub-cortical structures, higher mental processes were represented diffusely in the brain, all working as one.Gall and Flourens were vehemently opposed to each other’s work and ideas. Gall noted that Flourens’s procedure ‘mutilates all the organs at once, weakens them all, extirpates them all at the same time’. In fact, ablation by successive slices was a poor technique for demonstrating cortical localization and he was rightly reluctant to deduce general conclusions from lower organisms to primates and man. Flourens [8] attacked Gall’s efforts at localization and in contrast founded his theory of cerebral homogeneity that anticipated Lashley’s [9] much later concept of mass action and cortical equipotentiality. However, Flourens did concede that Gall had adduced detailed evidence of correlation between function and structure within the brain, and had established that the brain was the organ of mind. This was to be Gall’s most important contribution.Further DevelopmentsClinical localization was propounded by Jean-BaptisteBouillaud (1796–1881) [10] and Auburtin [11]. The psychological studies both of Alexander Bain and Herbert Spencer influenced the many advances of Hughlings Jackson.Precise localization remained, however, in dispute. On 4 April 1861, the Sociétéd’Anthropologie held a meeting. Paul Broca [12, 13] provided his now famous communication, which related partial destruction of the left frontal lobe to aphasia (aphemia) [14]. Jackson developed the concept that motor and sensory functions must be represented in an organized, hierarchical fashion. Fritsch and Hitzig, in 1870, showed in the dog that circumscribed cortical areas controlled movements of the contralateral limbs and that ablation caused weakness. Their findings confirmed the clinical localization of motor function. Testing Jackson’s notions, David Ferrier [15] used both ablation and electrical stimulation to produce topographical localization of function in several species. Published in his acclaimed The Functions of the Brain, Ferrier [16] had thereby mapped sensory and motor areas across several species, thus expanding both understanding and localization of movement and sensation [17].A celebrated debate in 1875 at the Société de Biologie in Paris between Charcot and Brown-Séquard illustrated the controversies of the day [18]. Charcot held an anatomical approach to localization of function. He relied on human pathological material primarily from isolated cerebral hematomas to establish the role of the cortex and subcortical white matter fibre tracts to motor and sensory function. By contrast, Brown-Séquard held a physiologist’s opinions, which disagreed fundamentally, arguing that since the brain consisted of complex networks, isolated lesions had no direct bearing on the localization of cerebral function. Though Brown-Séquard’s approach was to be vital in the evolving 20th-century concepts of brain function, it is generally agreed that in 1875, Charcot won the debate.In 1909, KorbinianBrodmann [19], stimulated by Oskar Vogt, described the different cytoarchitectonic structure of the gyri and showed that the human cortex is organized anatomically in the same way in man and other mammals. Brodmann’s discrete cortical areas [20] were, however, scorned with arguable justification by Henry Head. Other ContributionsFlourens [21] made other discoveries. In 1825 he observed that when a pigeon’s horizontal semi-circular canal was destroyed, the bird continued to turn in a circle, thus demonstrating the function of the vestibular labyrinth. He also showed that since destroying the medulla oblongata caused death of his animals, the medulla was responsible for circulation and breathing. In 1847, he described the anaesthetic properties of ether [22].Amongst Flourens’s pupils were EdméFélix Alfred Vulpian (1826–1887) and Gabriel Gustav Valentin (1810–1883). In 1840 he was elected to the French Academy, in preference to Victor Hugo. In 1845 he was created a commander of the légiond’honneur, and in the following year a peer of France. With the revolution in 1848 he withdrew from political life and in 1855 accepted the Professorship of Natural History at the Collège de France. He died at Montgeron, near Paris, on 6 December 1867.
- Bouillaud was born in 1796, studied medicine under Dupuytren and Magendie, being inspired by the latter with a profound interest in physiology. He received his doctorate in i823, and at the age of twentynine, published a brilliantmonograph, entitled "Traite clinique et physiologique de l'encephalite." In this volume he presented a number of clinical and pathological studies on loss of articulate speech associated with lesions of the anterior lobes of the brain, giving his reasons for localization of this function of the brain. In May of the same year, a further contribution was made, demonstrating that the loss of articulate speech corresponded to a lesion of the anterior lobes of the brain. These astute clinical studies won for him election to the Academy of Medicine in i826, at the age of thirty. Four years later, he became Professor of Medicine, succeeding Recamier, joining the staff of the Charit6, in i848, becoming Dean of the Faculty of Medicine, and in i862 President of the Academy of Medicine.Bouillaud described cases from the literature and from his own practice showing that those who had lost the faculty of speech had lesions in the anterior lobes at postmortem. He made it clear that speech could be lost as an isolated functional defect and that loss of speech was not due to inability to move the tongue or lips, but to a lesion in the anterior lobes of the brain. Bouillaud's conclusions that the center for the faculty of speech resides in these lobes were based on clinical and pathological grounds.In support of his view, Bouillaud offered evidence showing that involvement of the anterior lobes alone was associated with loss of the faculty of speech. These he termed "positive arguments." Others were presented in which the disease affected other parts of the brain and in which speech was not affected. These he called "negative arguments." He reasoned that: "If it is true that the anterior lobes preside over speech, how can this be established since animals do not possess speech and therefore physiological experiments, on animals which have so widely contributedto science could furnish no positive evidence. In fact, man is the only animal who enjoys the noble privilege of speech; yet, man is sacred and inviolable for experimental physiology. Yet, disease, nature's own experiment on man, no less ingenious than the most skillful physiologist,can furnish the elements essential for the solution of our problem.How often nature, by means of illness, destroys the functions of the anterior lobes of the brain, the kind of a lesion which for our end is equivalent to ablation itself." "If on the one hand physiology is the true light of pathology, it is equally certain that pathology is for physiology an inexhaustible source of information. It leads us in a most direct manner to an understanding of the function of organs.“Bouillaud held that "the frontal lobes, in effect, preside over the great act of speech; but it is not by speech alone and the operations whichaccompany it, such as writing, arithmetic, etc., that man exercises supreme control over his environment, and extends indefinitely therange of his intelligence and the sphere of his perfectability. It is, therefore, probable that the anterior part of the brain is not exclusivelydevoted to the important functions of articulate language, but that it is also the seat of the highest functions of judgment. Let us -then ceaseto be astonished if the frontal area of man proves an extent proportionately so superior to the frontal region of all other animals; it couldnot be otherwise since in man, in that part of the brain corresponding to the frontal lobes, are situated the centres whose sublime gift he alonehas received from nature.“Thus, not only does he place speech in the frontal lobes, but also the higher intellectual faculties. These revolutionary views concerningthe functions of the anterior lobes, he repeatedly presented during the ensuing years, before the Academy of Medicine and the Academy ofScience, based on both clinical and pathological material. Yet, in spite of the evidence offered, his views met with little acceptance.By his critical analyses of the clinical problems before him Bouillaud felt that the time had come when medicine could propose the followingproblem: "Given the symptoms, determine the seat of the disease," and conversely: "We shall see in fact, that symptoms vary according to thelocation of the cerebral affection; that muscular paralysis, for example, varies according to the location the lesion occupies, whether the anterior,middle or posterior lobes of the brain, and we shall by this method be able to determine some of the functions devolved upon various partsof which the cerebrum is composed. From this last point of view one of the most interesting results, to which clinical observations have led us,is that the anterior part of the brain is truly the centre of articulated language.“Bouillaud reasoned further that a focal and limited paralysis must indicate a focal and limited lesion of the brain, stating that: "The pluralityof cerebral centres reserved for movement is in effect proven by the existence of isolated partial paralysis corresponding to a local alterationof the brain; since it is evident that if this organ were not made up of several motor or conductor centres for muscular movement, it wouldbe impossible to conceive how a lesion of one of these areas carried with it a paralysis of a given part of the body without affecting movementof any of the other parts.“Bouillaud's contributions were not limited to the function of the cerebrum. In i 827 he published the results of his experimental andclinical studies indicating that the cerebellum controlled equilibrium and station, refuting Gall's view that the cerebellum was an organ ofthe instinct of propagation, thus recognizing important functions of the cerebellum.Thus, the basic principles overthrowing the concept that the brain acted as a whole were solidly laid by Bouillaud in i 825, and on numeroussubsequent occasions, notably in i839 and in 1848. Unfortunately, his monumental contributions in the field of neuro-anatomy and clinicalneurology have not even yet been generally recognized; whereas, his work on the heart and the relationship of rheumatic fever to endocarditishave been generally appreciated.*The opposition with which Bouillaud's views on cerebral localization were met seems hard to understand since they were adequatelysupported by clinical and pathological evidence; yet in spite of these clinical and pathological contributions indicating the presence of variouscenters in the brain for specific and localized functions the problem remained unsettled. We find a great debate concerning cerebral localizationand the existence of cerebral centers being carried on in the Anthropological Society of Paris during i86i, some thirty-six years after Bouillaud's original contributions.
- During February, March, and April, 1861, a notable series of meetings of the Anthropological Society took place in Paris, in which * At Bouillaud's request, no oration was held at his funeral; however, four years later his compatriots, recognizing his many contributions and the dominant role which he had played in medicine, erected a statue of him au Place Angouleme.Pierre Gratiolet, Ernest Auburtin and Paul Broca participated, which culminated finally in the recognition and establishment of the principleof cerebral localization.This was a great debate, a continuing debate, carried on primarily by Pierre Gratiolet, the fame of whose contribution in anatomy carriedsuch tremendous weight that it made any opposing opinion difficult to sustain. Nevertheless, undaunted, Ernest Auburtin, thirty-six yearsold, Chief of Clinic at Charite, devoted pupil and son-in-law of Bouillaud, repeatedly rose in the various meetings to disagree with Gratiolet'sconcept that the brain acted as a whole and that localized functional centers did not exist. Auburtin insisted that were one single localizedcerebral function established, then the principle of cerebral localization would be demonstrated. Were the principle of cerebral centers provenfor one function, the centers controlling other functions could then be unravelled and the brain elevated to its proper position, no longer conceivedof as acting as a single unit, as it had for centuries been thought to do.Auburtin held that if it can be demonstrated that lesions which abolish speech constantly occupy a definite cerebral convolution onecan hardly fail to admit that this convolution is the seat of articulated language. Proof of the existence of such a center of localized functionwould become the key to unlock additional centers.Gratiolet's paper at these meetings had to do with the size, volume, etc., of the brain as a whole from an anthropological viewpoint. Hepresented a Tolenac skull and indicated the importance of the volume of the brain, holding that size and weight were the determining factorsin intelligence and firmly expressing his belief that the brain acted as a whole, that all parts of the brain participated simultaneously andequally in intelligence.Broca also participated in these meetings. His paper was entitled "On the Volume and Form of the Brain According to Individuals andAccording to Race," thus carrying on in general the concept of Gratiolet that the size and shape of the brain determined its function as a whole.However, Broca felt that separation of the brain into anatomical divisions also meant separation of function, holding that we already hadenough evidence to say that all cerebral convolutions do not have the same attributes, maintaining that in the brain, as pointed out by Galland Spurzheim, there are distinct functions associated with anatomical division. Broca, however, while leaning toward the concept of cerebrallocalization did not come to the point of identifying any particular function with any particular part of the brain.Auburtin, opposing the views of Gratiolet, insisted that the opinion first expressed by Gall, and subsequently elaborated by Bouillaud,namely that the faculty of speech was to be found in the anterior lobe had been adequately demonstrated clinically. Auburtin agreed that thebrain was a complex organ with multiple and diverse functions but that greater advances would be made, and more precise information obtained,were the brain considered in its various parts, rather than as a whole.Auburtin argued, as did Bouillaud before him, that the highest cerebral functions reside in the anterior lobes and are in proportion to the development of these lobes. To this Gratiolet replied that research thus far had failed to find any localization of faculties in the brain. However,Auburtin called attention to the fact that various brain lesions produced quite different signs, depending upon their localization. The lesions ofthe anterior lobe produce neither loss of sensation nor of motion, yet abolish the faculty of speech. Gratiolet held that the difference dependedupon whether the lesion occurred slowly or rapidly. To this, Auburtin answered that while different pathological lesions may produce differentresults, it is none the less important that similar lesions in simikir areas produce similar results.Auburtin insisted on the importance of examining a particular function of the anterior lobes in order that the problem might be broughtinto sharper focus. He then called attention to a number of cases cited in the literature, notably those by Lallerman, Rosten, and particularlythose in Bouillaud's "Traite de l'encephalite," in which the clinical findings of loss of the faculty of articulate speech had permitted them topredicate and subsequently demonstrate at autopsy a lesion in the frontal lobes. Auburtin then reported a most striking clinical observation andclinical experiment which has been curiously neglected as one of the first instances of determining physiological data upon the human brainwith the patient conscious and able to cooperate in the experiment. A patient who had tried to commit suicide by shooting himself throughthe frontal region, entered l'Hopital St. Louis. A part of the frontal bone was removed, exposing the frontal lobe which was not lacerated. Thepatient's intelligence was intact, as well as his speech. He survived sufficiently long to be the subject of a very interesting experiment. When aspatula was applied to the anterior lobe, gently without causing general pressure, speech would instantly be abolished, only to return as soon asthe spatula was removed. This was done repeatedly, with repeated loss of the faculty of speech unaccompanied by any other paralysis. Its clinicalimplications and its significance were realized by Auburtin.Auburtin recognized that the anterior lobes were large and that, as yet, the precise area where the faculty of speech resides had not beendetermined. Instances could be cited in which a lesion existed in one anterior lobe and not the other, suggesting that perhaps one being intact,the intact lobe is able to contribute in part to the function of the other lobe.In order to bring the discussion which had taken place at length to a final conclusion, Auburtin on April 4, challenged those who opposedcerebral localization by saying that "On the service of M. Bouillaud, I have studied for a long time a patient named Bache who has lost hisspeech, who, nevertheless understands all that is said to him, replying by signs in an intelligent manner to all questions put to him. This patienthas spent several years at Bicetre and is now at the Hospital for Incurables.I have seen him recently, his illness has progressed, a slight paralysis has supervened, but his intelligence is still intact and his speech stillabolished. This man will die without doubt, in a short time. In view of the symptoms which he presents, I have made a diagnosis of softeningof the anterior lobes. If at autopsy the anterior lobes are found intact, then I shall renounce the ideas which I have sustained, but I can onlyreason in accordance with the facts which today exist in science. Still, I have never seen a lesion limited to the middle or posterior lobe destroythe faculty of speech.“To this Gratiolet replied that he could cite cases in which the frontal lobes were involved without loss of speech and replied: "I do not hesitateto conclude that all attempts at localization, which up to now have been tried, lack any foundation. They are no doubt great efforts, titanicefforts! But when one attempts to grasp the truth at the height of these babbles, the edifice crumbles." It is noteworthy that throughout thisdebate in which Gratiolet did not hesitate to be emphatic in his views concerning the fallacy of cerebral localization and Auburtin equallyvigorously attacking the concept that the brain acted as a whole, as a single unit, Broca though present at each of the sessions did not commithimself as to the fundamental concept concerning the relationship of the faculty of speech and the anterior lobes.
- Electrically stimulated various parts of a dog's motor cortex. They observed that depending on what part of the cortex they stimulated, a different part of the body contracted. Then they found that if they destroyed this same small area of the cortex, the corresponding part of the body became paralyzed. This is how it was discovered that every part of the body has a particular region of the primary motor cortex that controls its movement
- David FerrierIn 1870, two German scientists, Gustav Fritsch and Eduard Hitzig, reported their findings on the electrical excitability of the exposed cerebral cortex in dogs in which galvanic current was used. According to Horwitz,[5] David Ferrier was impressed by these scientists' work, and he refined their techniques. By using faradic current stimulation, Ferrier investigated even further the cortical functioning of many different animals, including primates. He constructed one of the first detailed cortical maps and confirmed many of the principles set forth by Dr. Jackson. Ferrier firmly established the location of the motor cortex, stating that it extended along the rolandic fissure medially to the interhemispheric area. Not all of his conclusions were correct, however. For example, he erroneously placed the center for vision in the superior temporal lobes, and similarly was unable adequately to determine the function of the occipital lobes from his studies. Ferrier summarized his results in the 1876 publication of The Functions of the Brain ,[4] and dedicated this work to Jackson, stating "To Dr. Hughlings Jackson, who from a clinical and pathological standpoint anticipated many of the more important results of recent experimental investigation into the functions of the cerebral hemispheres." As discussed in Horwitz,[5] 2 years later, Ferrier produced a second publication titled The Localisation of Cerebral Disease , and he dedicated it to a well-known French neurologist, Dr. Jean-Martin Charcot.
- John Hughlings JacksonJohn Hughlings Jackson, one of the greatest early contributors to the current knowledge of anatomical cerebral localization, was born in England in 1835. After receiving his medical education and initial training in York, he came to London in 1859. There he was substantially influenced by an acquaintance, Dr. Charles Brown-Séquard, to pursue his interest in the study of neurology, particularly seizures. Jackson, an astute clinician, obtained finely detailed histories from and performed thorough examinations on his patients. Then, with tireless attention to minutiae, he observed the natural and clinical courses of diseases and carefully correlated these findings with the pathological findings at autopsy (see Horwitz).[7]Until Jackson's time, seizures were thought to originate in the medulla oblongata. According to Horwitz,[7] however, based on extensive study of patients with epilepsy, Jackson concluded that an abnormal electrical discharge from the cortical gray matter was consistently the origin of the seizures in his patients. He described individuals who had sustained a blunt injury to the head and in whom hemiplegia subsequently developed on the contralateral side of the body and face. When these patients experienced epileptic activity, with rare exceptions it began on the hemiplegic side. Jackson[9] also noted that those whose external injury overlaid the posterior left frontal lobe usually had a degree of difficulty with speech at baseline or during their seizure. Extrapolating this information to other atraumatic scenarios, he was often able to predict with great accuracy the location of intracerebral lesions found at autopsy. The following excerpt is taken from "A Study of Convulsions," which appeared in the publication Transactions St. Andrews Medical Graduates , volume iii, in 1870 (see Horwitz).[7]The great majority of chronic convulsions may be arranged in two classes.Those in which the spasm affects both sides of the body almost contemporaneously.Those in which the fit begins by deliberate spasm on one side of the body, and in which parts of the body are affected, one after another.It is with the second class only that I intend to deal in the article...Fits beginning unilaterally may doubtless begin by movement in any part of the region which is paralysed in hemiplegia, i.e. in the face, in the arm, or in the leg. But I know few casesof fits of this class which begin other than in the side of the face (usually the cheek), in the hand, or in the foot... The fit usually begins, it is to be observed, in that part of the face, of the arm, and of the leg which has the most varied uses.Parts with the most varied uses will be represented in the central nervous system by most ganglion cells. I say most varied movements, as it is not only a question of number of movements, but also of number of different movements.The eponym "jacksonian seizure" or "jacksonian march" was derived from the foregoing description.[7]
- Sir Charles Scott Sherrington 1857-1952, English neurophysiologist, educated at Cambridge. He was professor of physiology at the universities of Liverpool and London and at Oxford. He contributed major concepts in his field, among them that of proprioception, that of the function of the synapse (a term he introduced), and the process described in his Integrative Action of the Nervous System (1906, 2d ed. 1948). As a physician, he did important work in the study of cholera and of diphtheria and tetanus antitoxins, and played an important role in the improvement of health and safety conditions in British factories during World War I. He was knighted in 1922 and with E. D. Adrian shared the 1932 Nobel Prize in Physiology or Medicine for their discoveries regarding the function of the neuron. Among his other works are Mammalian Physiology (1919, rev. ed. 1929), The Brain and Its
- Commands for voluntary movement originate in cortical association areas. The movements are planned in the cortex as well as in the basal ganglia and the lateral portions of the cerebellar hemispheres, as indicated by increased electrical activity before the movement. The basal ganglia and cerebellum both funnel information to the premotor and motor cortex by way of the thalamus. Motor commands from the motor cortex are relayed in large part via the corticospinal tracts to the spinal cord and the corresponding corticobulbar tracts to motor neurons in the brain stem. However, collaterals from these pathways and a few direct connections from the motor cortex end on brain stem nuclei, which also project to motor neurons in the brain stem and spinal cord. These pathways can also mediate voluntary movement. Movement sets up alterations in sensory input from the special senses and from muscles, tendons, joints, and the skin. This feedback information, which adjusts and smoothes movement, is relayed directly to the motor cortex and to the spinocerebellum. The spinocerebellum projects in turn to the brain stem. The main brain stem pathways that are concerned with posture and coordination are the rubrospinal, reticulospinal, tectospinal, and vestibulospinal tracts and corresponding projections to motor neurons in the brain stem. Ganong 21st
- Spinal cord reflex mechanisms involve lower motor neurons and are subject to supraspinal control. The brain regions that have direct control over the spinal cord include the brainstem and cerebral cortex. Brainstem pathways that project to the spinal cord include the reticulospinal, vestibulospinal, rubrospinal, and tectospinal tracts. The cerebral cortex gives rise to both corticospinal and corticobulbar fibers. Corticospinal fibers are essential for voluntary control over fine movements, mainly of the distal extremities. Corticobulbar fibers contribute to the control of spinal cord indirectly, by acting on neurons of the brainstem that project to the spinal cord. Other corticobulbar fibers innervate lower motor neurons of the brainstem (cranial nerves) and provide the substrate and mechanism for voluntary movements of the head region. Two other regions, the basal ganglia and cerebellum, play important roles in motor functions. The basal ganglia affect motor systems by acting on neurons in the precentral and premotor regions that comprise the larger part of the corticospinal tract. The cerebellum affects motor function by acting on neurons in both the brainstem and the cerebral cortex that directly control motor functions of the spinal cord.
- Figure 12-3. Medial (above) and lateral (below) views of the human cerebral cortex, showing the motor cortex (Brodmann's area 4) and other areas concerned with control of voluntary movement, along with the numbers assigned to the regions by Brodmann. (Reproduced, with permission, from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) By means of stimulation experiments in patients undergoing craniotomy under local anesthesia, it has been possible to outline most of the motor projections from the motor cortex. These have been confirmed in unanesthetizedunoperated humans by PET scanning and fMRI (Figure 12-4). The various parts of the body are represented in the precentralgyrus, with the feet at the top of the gyrus and the face at the bottom (Figure 12-5). The facial area is represented bilaterally, but the rest of the representation is unilateral, the cortical motor area controlling the musculature on the opposite side of the body. The cortical representation of each body part is proportionate in size to the skill with which the part is used in fine, voluntary movement. The areas involved in speech and hand movements are especially large in the cortex; use of the pharynx, lips, and tongue to form words and of the fingers and apposable thumbs to manipulate the environment are activities in which humans are especially skilled. The conditions under which the human stimulation studies were performed precluded stimulation of the banks of the sulci and other inaccessible areas. Meticulous study has shown that in monkeys, there is a regular representation of the body, with the axial musculature and the proximal portions of the limbs represented along the anterior edge of the precentralgyrus and the distal part of the limbs along the posterior edge. Another feature of M1 is the presence of considerable overlap in the muscles that are innervated, as well as innervation of synergic muscles separated by considerable distances. There has been debate about whether individual muscles or movements are represented in M1, and the most recent evidence indicates that both are represented. The cells in the cortical motor areas are arranged in columns. The cells in each column receive fairly extensive sensory input from the peripheral area in which they produce movement, providing the basis for feedback control of movement. Some of this input may be direct, and some is relayed from somatic sensory area I in the postcentralgyrus. Cerebral dominance, which is discussed in detail in Chapter 16, also affects the motor cortex in humans. Moving the fingers of the left hand is associated mainly with activation of the right motor cortex and vice versa, as measured by imaging techniques (see Chapter 32). However, moving the fingers of the left hand also activates the left motor cortex, particularly in individuals who are right-handed. This correlates with the fact that lesions of the left motor cortex cause motor dysfunction in the left hand as well as the right hand, whereas lesions of the right motor cortex have little effect on the right hand. Supplementary Motor AreaFor the most part, the supplementary motor area projects to the motor cortex. It appears to be involved primarily in programming motor sequences. Lesions of this area in monkeys produce awkwardness in performing complex activities and difficulty with bimanual coordination. When human subjects count to themselves without speaking, the motor cortex is quiescent, but when they speak the numbers aloud as they count, blood flow increases in the motor cortex and the supplementary motor area. Thus, the supplementary motor area as well as the motor cortex is involved in voluntary movement when the movements being performed are complex and involve planning. Blood flow increases whether or not a planned movement is carried out. The increase occurs whether the movement is performed by the contralateral or the ipsilateral hand. Premotor CortexThe premotor cortex projects to the brain stem areas concerned with postural control and to the motor cortex as well as providing part of the corticospinal and corticobulbar output. Its function is still incompletely understood, but it may be concerned with setting posture at the start of a planned movement and with getting the individual ready to perform. Posterior Parietal CortexIn addition to providing fibers that run in the corticospinal and corticobulbar tracts, the somatic sensory area and related portions of the posterior parietal lobe project to the premotor area. Lesions of the somatic sensory area cause defects in motor performance that are characterized by inability to execute learned sequences of movements such as eating with a knife and fork. Some of the neurons in area 5 (Figure 12-3) are concerned with aiming the hands toward an object and manipulating it, whereas some of the neurons in area 7 are concerned with hand-eye coordination. Role in MovementThe corticospinal and corticobulbar system is the primary pathway for the initiation of skilled voluntary movement. This does not mean that movement—even skilled movement—is impossible without it. Nonmammalian vertebrates have essentially no corticospinal and corticobulbar system, but they move with great agility. Cats and dogs stand, walk, run, and even eat if food is presented to them after complete destruction of this system. Only in primates are relatively marked deficits produced. Careful section of the pyramids producing highly selective destruction of the lateral corticospinal tract in laboratory primates produces prompt and sustained loss of the ability to grasp small objects between two fingers and to make isolated movements of the wrists. However, the animal can still use the hand in a gross fashion and can stand and walk. These deficits are consistent with loss of control of the distal musculature of the limbs, which is concerned with fine skilled movements. On the other hand, lesions of the ventral corticospinal tract produce axial muscle deficits that cause difficulty with balance, walking, and climbing. Ganong 21stPrimary Motor CortexNote that more than one half of the entire primary motor cortex is concerned with controlling the muscles of the hands and the muscles of speech. Point stimulation in these hand and speech motor areas on rare occasion causes contraction of a singlemuscle; most often, stimulation contracts a group of muscles instead. To express this in another way, excitation of a single motor cortex neuron usually excites a specific movement rather than one specific muscle.To do this, it excites a “pattern” of separate muscles, each of which contributes its own direction and strength of muscle movement.Premotor AreaThe premotor area, also shown in Figure 55–1, lies 1 to 3 centimeters anterior to the primary motor cortex, extending inferiorly into the sylvian fissure and superiorly into the longitudinal fissure, where it abuts the supplementary motor area, which has functions similar to those of the premotor area. The topographical organization of the premotor cortex is roughly the same as that of the primary motor cortex, with themouth and face areas located most laterally; as one moves upward, the hand, arm, trunk, and leg areas are encountered.Nerve signals generated in the premotor area cause much more complex “patterns” of movement than the discrete patterns generated in the primary motor cortex. For instance, the pattern may be to position the shoulders and arms so that the hands are properly oriented to perform specific tasks. To achieve these results, the most anterior part of the premotor area first develops a “motor image” of the total musclemovement that is to be performed.Then, in the posterior premotor cortex, this image excites each successive pattern of muscle activity required to achieve the image.This posterior part of the premotor cortex sends its signals either directly to the primary motor cortex to excite specific muscles or, often, by way of the basal ganglia and thalamus back to the primary motor cortex. Thus, the premotor cortex, basal ganglia, thalamus,and primary motor cortex constitute a complex overall system for the control of complex patterns of coordinated muscle activity.Supplementary Motor AreaThe supplementary motor area has yet another topographical organization for the control of motor function. It lies mainly in the longitudinal fissure but extends a few centimeters onto the superior frontal cortex. Contractions elicited by stimulating this area are often bilateral rather than unilateral. For instance, stimulation frequently leads to bilateral grasping movements of both hands simultaneously; thesemovements are perhaps rudiments of the hand functions required for climbing. In general, this area functions in concert with the premotor area to provide body-wide attitudinal movements, fixation movements of the different segments of the body, positional movements of the head and eyes, and so forth, as background for the finer motor control of the arms and hands by the premotor area and primary motor cortex.
- Some Specialized Areas of Motor Control Found in the Human Motor CortexNeurosurgeons have found a few highly specialized motor regions of the human cerebral cortex (shown in Figure 55–3) that control specific motor functions.These regions have been localized either by electricalstimulation or by noting the loss of motor functionwhen destructive lesions occur in specific corticalareas. Some of the more important regions are thefollowing.Broca’s Area and Speech. Figure 55–3 shows a premotorarea labeled “word formation” lying immediatelyanterior to the primary motor cortex and immediatelyabove the sylvian fissure. This region is called Broca’sarea. Damage to it does not prevent a person fromvocalizing, but it does make it impossible for theperson to speak whole words rather than uncoordinatedutterances or an occasional simple word such as“no” or “yes.” A closely associated cortical area alsocauses appropriate respiratory function, so that respiratoryactivation of the vocal cords can occur simultaneouslywith the movements of the mouth and tongueduring speech. Thus, the premotor neuronal activitiesrelated to speech are highly complex.“Voluntary” Eye Movement Field. In the premotor areaimmediately above Broca’s area is a locus for controllingvoluntary eye movements. Damage to this areaprevents a person from voluntarily moving the eyestoward different objects. Instead, the eyes tend to lockinvoluntarily onto specific objects, an effect controlledby signals from the occipital visual cortex, as explainedin Chapter 51. This frontal area also controls eyelidmovements such as blinking.Head Rotation Area. Slightly higher in the motor associationarea, electrical stimulation elicits head rotation.This area is closely associated with the eye movementfield; it directs the head toward different objects.Area for Hand Skills. In the premotor area immediatelyanterior to the primary motor cortex for the hands andfingers is a region neurosurgeons have identified asimportant for “hand skills.” That is, when tumors orother lesions cause destruction in this area, handmovements become uncoordinated and nonpurposeful,a condition called motor apraxia.
- Red Nucleus Serves as an Alternative Pathway for Transmitting Cortical Signals to the Spinal CordThe red nucleus, located in the mesencephalon, functions in close association with the corticospinal tract.As shown in Figure 55–5, it receives a large number of direct fibers from the primary motor cortex through the corticorubral tract, as well as branching fibers from the corticospinal tract as it passes through the mesencephalon.These fibers synapse in the lower portion of the red nucleus, the magnocellular portion, which contains large neurons similar in size to the Betz cells in the motor cortex.These large neurons then give rise to the rubrospinal tract, which crosses to the opposite side in the lower brain stem and follows a course immediately adjacent and anterior to the corticospinal tract into the lateral columns of the spinal cord.The rubrospinal fibers terminate mostly on the interneurons of the intermediate areas of the cord gray matter, along with the corticospinal fibers, but some of the rubrospinal fibers terminate directly on anterior motor neurons, along with some corticospinal fibers.The red nucleus also has close connections with the cerebellum, similar to the connections between the motor cortex and the cerebellum.Function of the Corticorubrospinal System. The magnocellularportion of the red nucleus has a somatographic representation of all the muscles of the body, as is true of the motor cortex. Therefore, stimulation of a single point in this portion of the red nucleus causes contraction of either a single muscle or a small group of muscles. However, the fineness of representation of the different muscles is far less developed than in the motor cortex. This is especially true in human beings,who have relatively small red nuclei.The corticorubrospinal pathway serves as an accessory route for transmission of relatively discrete signals from the motor cortex to the spinal cord.When the corticospinal fibers are destroyed but the corticorubrospinal pathway is intact, discrete movements can still occur, except that the movements for fine control of the fingers and hands are considerably impaired. Wrist movements are still functional, which is not the case when the corticorubrospinal pathway is also blocked.Therefore, the pathway through the red nucleus to the spinal cord is associated with the corticospinal system. Further, the rubrospinal tract lies in the lateral columns of the spinal cord, along with the corticospinal tract, and terminates on the interneurons and motor neurons that control the more distal muscles of the limbs.Therefore, the corticospinal and rubrospinal tracts together are called the lateral motor system of the cord, in contradistinction to a vestibuloreticulospinalsystem, which lies mainly medially in the cord and is called the medial motor system of the cord, asdiscussed later in this chapter.
- Muscle SpindlesMuscle spindles are small encapsulated sensory receptors that have a spindle-like or fusiform shape and are located within the fleshy part of the muscle. Their main function is to signal changes in the length of the muscle within which they reside. Changes in the length of muscles are closely associated with changes in the angles of the joints that the muscles cross. Thus, muscle spindles can be used by the central nervous system to sense relative positions of the body segments.Each spindle has three main components: (1) a group of specialized intrafusalmuscle fibers whose central regions are noncontractile; (2) large-diameter myelinated sensory endings that originate from the central regions of the intrafusal fibers; and (3) small-diameter myelinated motor endings that innervate the polar contractile regions of the intrafusal fibers (Figure 36-3A). When the intrafusal fibers are stretched, often referred to as “loading the spindle,” the sensory endings are also stretched and increase their firing rate. Because muscle spindles are arranged in parallel with the extrafusal muscle fibers that make up the main body of the muscle, the intrafusal fibers change in length as the whole muscle changes. Thus, when a muscle is stretched, the activity in the sensory endings of muscle spindles is increased. When a muscle shortens, the spindle is unloaded and the activity decreases.The motor innervation of the intrafusal muscle fibers comes from small-diameter motor neurons, called gamma motor neurons to distinguish them from the large-diameter alpha motor neurons that innervate the extrafusal muscle fibers. Contraction of the intrafusal muscle fibers does not contribute to the force of muscle contraction. Rather, activation of gamma motor neurons causes shortening of the polar regions of the intrafusal fibers. This in turn stretches the noncontractile central region from both ends, leading to an increase in firing rate of the sensory endings or to a greater likelihood that stretch of the muscle will cause the sensory ending to fire. Thus, the gamma motor neurons provide a mechanism for adjusting the sensitivity of the muscle spindles.The structure and functional behavior of muscle spindles is considerably more complicated than this simple description implies. When a muscle is stretched, there are two phases of the change in length: a dynamic phase, the period during which length is changing, and a static or steady-state phase, when the muscle has stabilized at a new length. Structural specializations within each component of the muscle spindles allow spindle afferents to signal aspects of each phase separately.There are two types of intrafusal muscle fibers: nuclear bag fibers and nuclear chain fibers. The bag fibers can be divided into two groups, dynamic and static. A typical spindle has 2 or 3 bag fibers and a variable number of chain fibers, usually about 5. Furthermore, there are two types of sensory fiber endings: a single primary ending and a variable number of secondary endings (up to 8). The primary (Ia fiber) ending spirals around the central region of all the intrafusal muscle fibers (Figure 36-3B). The secondary (group II fiber) endings are located adjacent to the central regions of the static bag and chain fibers. The gamma motor neurons can also be divided into two classes, dynamic and static. Dynamic gamma motor neurons innervate the dynamic bag fibers, while the static gamma motor neurons innervate the static bag and the chain fibers.This duality of structure is reflected in a duality of function. The steady-state or tonic discharge of both primary and secondary sensory endings signals the steady-state length of the muscle. The primary endings are, in addition, highly sensitive to the velocity of stretch, allowing them to provide information about the speed of movements. Because they are highly sensitive to small changes, primary endings provide quick information about unexpected changes in length, useful for generating quick corrective reactions.Increases in activity of dynamic gamma motor neurons increase the dynamic sensitivity of the primary endings but have no influence on the secondary endings. Increases in activity of static gamma motor neurons increase the tonic level of activity in both primary and secondary endings, decrease the dynamic sensitivity of primary endings, and can prevent the silencing of primary activity when a muscle is released from stretch (Figure 36-3C). Thus, the central nervous system can independently adjust the dynamic and static sensitivity of the sensory fibers from muscle spindles.Box 36-2 Selective Activation of Sensory Fibers from MuscleSensory fibers are classified according to their diameter. Axons with larger diameters conduct action potentials more rapidly. Because each class of receptors gives rise to afferent fibers with diameters within a restricted range, this method of classification distinguishes to some extent the fibers that arise from the different groups of sensory receptors. The main groups of sensory fibers from muscle are listed in Table 36-1 (see Chapter 24 for the classification of sensory fibers from skin and joints).The organization of reflex pathways in the spinal cord has been established primarily by electrically stimulating the sensory fibers and recording evoked responses in different classes of neurons in the spinal cord. This method of activation has three advantages over natural stimulation. The timing of afferent input can be precisely established, the central responses evoked by different classes of sensory fibers can be assessed by grading the strength of the electrical stimulus, and certain classes of receptors can be activated in isolation (impossible in natural conditions).The strength of electrical stimuli required to activate a sensory fiber is measured relative to the strength required to activate the largest afferent fibers since the largest fibers have the lowest threshold for electrical activation. Thus group I fibers are usually activated over the range of one to two times the threshold of the largest afferents (with Ia fibers having, on average, a slightly lower threshold than Ib fibers). Most group II fibers are activated over the range of 2-5 times the threshold, while the small group III and IV fibers require stimulus strengths in the range of 10-50 times the threshold for activation.Reciprocal innervation of opposing muscles is not the only useful mode of coordination. Sometimes it is advantageous to contract the prime mover and the antagonist at the same time. Such co-contraction has the effect of stiffening the joint and is most useful when precision and joint stabilization are critical. An example of this phenomenon is the co-contraction of flexor and extensor muscles of the elbow immediately before catching a ball. The Ia inhibitory interneurons receive both excitatory and inhibitory signals from all of the major descending pathways (Figure 36-5A). By changing the balance of excitatory and inhibitory inputs onto these interneurons, supraspinal centers can reduce reciprocal inhibition and enable co-contraction, thus controlling the relative amount of joint stiffness to meet the requirements of the motor act.The activity of spinal motor neurons is also regulated by another important class of inhibitory interneurons, the Renshaw cells (Figure 36-5B). Renshaw cells are excited by collaterals of the axons of motor neurons, and they make inhibitory synaptic connections to several populations of motor neurons, including the same motor neurons that excite them, and to the Ia inhibitory interneurons. The connections of Renshaw cells to motor neurons form a negative feedback system that may help stabilize the firing rate of the motor neurons, while the connections to the Ia inhibitory interneurons may regulate the strength of reciprocal inhibition to antagonistic motor neurons. In addition, Renshaw cells receive significant synaptic input from descending pathways and distribute inhibition to task-related groups of motor neurons and Iainterneurons. Thus, it is likely that they contribute to establishing the pattern of transmission in divergent group Ia pathways according to the motor task.
- The Spinal Cord Circuitry Underlying Muscle Stretch ReflexesThe local circuitry within the spinal cord mediates a number of sensory motor reflex actions. The simplest of these reflex arcs entails the response to musclestretch, which provides direct excitatory feedback to the motor neurons innervating the muscle that has been stretched (Figure 16.9). As already mentioned, the sensory signal for the stretch reflex originates in muscle spindles, sensory receptors embedded within most muscles (see previous section and Chapter 9). The spindles comprise 8–10 intrafusal fibers arranged in parallel with the extrafusal fibers that make up the bulk of the muscle (Figure 16.9A). Large-diameter sensory fibers, called Ia afferents, are coiled around the central part of the spindle. These afferents are the largest axons in peripheral nerves and, since action potential conduction velocity is a direct function of axon diameter (see Chapters 2 and 3), they allow for very rapid adjustments in this reflex arc when the muscle is stretched. The stretch imposed on the muscle deforms the intrafusal muscle fibers, which in turn initiate action potentials by activating mechanically gated ion channels in the afferent axons coiled around the spindle. The centrally projecting branch of the sensory neuron forms monosynaptic excitatory connections with the α motor neurons in the ventral horn of the spinal cord that innervate the same (homonymous) muscle and, via local circuit neurons, inhibitory connections with the α motor neurons of antagonistic (heteronymous) muscles. This arrangement is an example of what is called reciprocal innervation and results in rapid contraction of the stretched muscle and simultaneous relaxation of the antagonist muscle. All of this leads to especially rapid and efficient responses to changes in the length or tension in the muscle (Figure 16.9B). The excitatory pathway from a spindle to the α motor neurons innervating the same muscle is unusual in that it is a monosynaptic reflex; in most cases, sensory neurons from the periphery do not contact the lower motor neuron directly but exert their effects through local circuit neurons.This monosynaptic reflex arc is variously referred to as the “stretch,” “deep tendon,” or “myotatic reflex,” and it is the basis of the knee, ankle, jaw, biceps, or triceps responses tested in a routine neurological examination. The tap of the reflex hammer on the tendon stretches the muscle and therefore excites a volley of activity from the muscle spindles in the afferent axons. The afferent volley is relayed to the α motor neurons in the brainstem or spinal cord, and an efferent volley returns to the muscle (see Figure 1.5). Since muscles are always under some degree of stretch, this reflex circuit is normally responsible for the steady level of tension in muscles called muscle tone. Changes in muscle tone occur in a variety of pathological conditions, and it is these changes that are assessed by examination of tendon reflexes.In terms of engineering principles, the stretch reflex arc is a negative feedback loop used to maintain muscle length at a desired value (Figure 16.9C). The appropriate muscle length is specified by the activity of descending pathways that influence the motor neuron pool. Deviations from the desired length are detected by the muscle spindles, since increases or decreases in the stretch of the intrafusal fibers alter the level of activity in the sensory fibers that innervate the spindles. These changes lead in turn to adjustments in the activity of the α motor neurons, returning the muscle to the desired length by contracting the stretched muscle and relaxing the opposed muscle group, and by restoring the level of spindle activity to what it was before.The smaller γ motor neurons control the functional characteristics of the muscle spindles by modulating their level of excitability. As already described, when the muscle is stretched, the spindle is also stretched and the rate of discharge in the afferent fibers increased. When the muscle shortens, however, the spindle is relieved of tension, or “unloaded,” and the sensory axons that innervate the spindle might therefore be expected to fall silent during contraction. However, they remain active. The γ motor neurons terminate on the contractile poles of the intrafusal fibers, and the activation of these neurons causes intrafusal fiber contraction—in this way maintaining the tension on the middle (or equatorial region) of the intrafusal fibers where the sensory axons terminate. Thus, co-activation of the α and γ motor neurons allows spindles to function (i.e., send information centrally) at all muscle lengths during movements and postural adjustments
- SummaryFour distinct but highly interactive motor subsystems—local circuits in the spinal cord and brainstem, descending upper motor neuron pathways that controlthese circuits, the basal ganglia, and the cerebellum—all make essential contributions to motor control. Alpha motor neurons located in the spinal cord and in the cranial nerve nuclei in the brainstem directly link the nervous system and muscles, with each motor neuron and its associated muscle fibers constituting a functional entity called the motor unit. Motor units vary in size, amount of tension produced, speed of contraction, and degree of fatigability. Graded increases in muscle tension are mediated by both the orderly recruitment of different types of motor units and an increase in motor neuron firing frequency. Local circuitry involving sensory inputs, local circuit neurons, and α and γ motor neurons are especially important in the reflexive control of muscle activity. The stretch reflex is a monosynaptic circuit with connections between sensory fibers arising from muscle spindles and the α motor neurons that innervate the same or synergistic muscles. Gamma motor neurons regulate the gain of the stretch reflex by adjusting the level of tension in the intrafusal muscle fibers of the muscle spindle. This mechanism sets the baseline level of activity in α motor neurons and helps to regulate muscle length and tone. Other reflex circuits provide feedback control of muscle tension and mediate essential functions such as the rapid withdrawal of limbs from painful stimuli. Much of the spatial coordination and timing of muscle activation required for complex rhythmic movements such as locomotion are provided by specialized local circuits called central pattern generators. Because of their essential role in all of these circuits, damage to lower motor neurons leads to paralysis of the associated muscle and to other changes, including the loss of reflex activity, the loss of muscle tone, and eventually muscle atrophy.