En 1903, lefrançais Victor Henri montre qu’une enzyme peut fixer un
ligand (tout ce qui se fixe sur une protéine est appelé ligand).
En 1913, Maud Menten introduit avec Leonor Michaelis un nouveau
concept qui révolutionne les études des réactions biologiques.
Notion de complexe enzyme-substrat
Complexe enzyme-substrat
ajout de
l’enzyme [E]T
onest au temps 0
[E]T pour [E]TOTALE
milieu réactionnel
contenant à t=0 une
concentration de
substrat [S]0
Complexe enzyme-substrat
à t=0, laconcentration en substrat est désignée [S]0
à t=0, la concentration en produit est égale à 0
la concentration en enzyme est désignée [E]T et reste
constante tout au long de la mesure
à t=0, la concentration du complexe enzyme-substrat, [ES],
est égale à 0
E + S E + P
ES
Complexe enzyme-substrat
mesure d’une activitéenzymatique
La concentration en enzyme doit rester constante au sein de tous les
essais.
Les paramètres physico-chimiques (température, force ionique et pH)
du milieu doivent rester stables pour tous les essais.
Sauf , le seul paramètre variable doit être la concentration en
substrat.
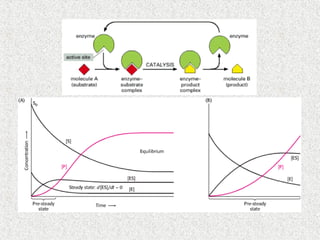
Phases de la réaction
10.
concentration en substrat[S]
vitesse
initiale
(Vi)
Vmax
[E]T cste
Équation d’une
hyperbole
Phases de la réaction
11.
E + SE + P
ES
k1
k-1
k2
vi = k2 [ES]
La vitesse d’apparition du produit P (vitesse de la
réaction catalysée par l’enzyme) dépend de k2 et de la
concentration en ES.
La [ES] dépend de sa vitesse de formation et de sa
vitesse de disparition.
vformation ES = k1 [E] [S]
vdisparition ES = k-1 [ES] + k2 [ES] = (k-1 + k2) [ES]
Équation de Michaelis-Menten
(k-1 + k2)[ES] = k1 [E] [S]
vformation ES = vdisparition ES
Pendant l'état dit "stationnaire" ou en "vitesse initiale", les
vitesses de formation et de disparition de ES sont égales.
Si on réarrange l’expression mathématique :
[ES] =
k1 [E] [S]
(k-1 + k2)
ou
[ES] =
[E] [S]
(k-1 + k2)
k1
Équation de Michaelis-Menten
14.
KM =
(k-1 +k2)
k1
On simplifie l’équation en définissant une nouvelle constante, KM,
appelée constante de Michaelis et Menten :
Si on réarrange l’expression mathématique :
[ES] =
[E] [S]
KM
Équation de Michaelis-Menten
[ES] =
[E] [S]
(k-1 + k2)
k1
15.
la [S] estégale à celle de départ ([S]0), puisque la concentration en
substrat est très largement supérieure à celle en enzyme.
[ES] =
[E] [S]
KM
La concentration en enzyme libre [E] doit être égale à la concentration
totale en enzyme ([E]T) moins la concentration en enzyme lié ([ES]).
[E] = [E]T – [ES]
[ES] =
[E] [S]
KM
devient [ES] =
([E]T – [ES]) [S]
KM
Équation de Michaelis-Menten
16.
[ES] =
([E]T –[ES]) [S]
KM
Si on développe
cela devient [ES] =
[E]T [S] – [ES] [S]
KM
[ES] + =
[E]T [S]
KM
[ES] [S]
KM
[ES] =
[S]
KM
[E]T
1 +
[S]
KM
[ES] ( 1 + ) =
[E]T [S]
KM
[S]
KM
Équation de Michaelis-Menten
17.
[ES] =
[S]
KM
[E]T
1 +
[S]
KM
Oron sait que : Vi = k2 [ES]
Donc on peut écrire : Vi = k2
[S]
KM
[E]T
1 +
[S]
KM
On peut simplifier :
vi = k2
[S]
KM
[E]T
+
[S]
KM
KM
KM
= k2
[E]T
[S]
[S]
KM +
Équation de Michaelis-Menten
18.
On sait quelorsque toutes les molécules d’enzyme sont saturées,
[ES] = [E]T et la vitesse de transformation du substrat en produit
est alors maximale :
Vmax = k2 [E]T
L’équation de Michaelis-Menten devient :
vi = Vmax
[S]
[S]
KM +
Vitesse
initiale
Équation de Michaelis-Menten
vi = k2
[E]T
[S]
[S]
KM +
19.
vi = Vmax
[S]
[S]
KM+
Si vi = ½ Vmax
½ Vmax = Vmax
[S]
[S]
KM +
½ (KM + [S]) = [S]
½ KM = [S] - ½ [S]
½ KM = ½ [S]
KM = [S]
KM est inversement proportionnelle à l’affinité de l’enzyme. Plus
l’affinité est élevée, et moins il faudra de substrat pour que
l’enzyme fonctionne.
Constante de Michaelis-Menten
KM est la concentration en substrat
pour laquelle l’enzyme fonctionne à
la moitié de sa vitesse maximale
20.
KM s’exprime enunité de concentration de substrat.
KM = [S] lorsque vi = ½ Vmax
mole/L
µmole/L
mg/L
g/L
etc…
Constante de Michaelis-Menten
Nomenclature :
M signifie Michaelis mais,
officiellement, il faudrait
écrire cette constante Km
21.
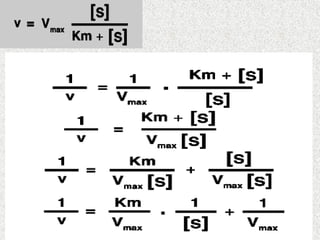
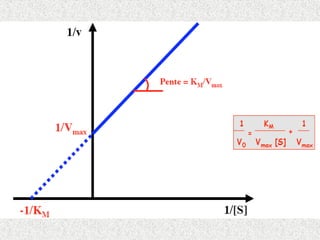
Détermination de Kmet Vmax:
Graphiquement on ne peut pas déterminer Km
et Vmax a partir de la courbe v= f [s]
On préfère placer les points expérimentaux sur
un diagramme ou ils s’alignent sur une droite.
Parmi les différentes transformation de
l’equation de michaelis réaliser dans ce but: la
représentation de lineweaver et burk (L.B)la
représentation en double inverse.
Consiste à porter les variation de 1/v en
fonction de 1/ [s]
24.
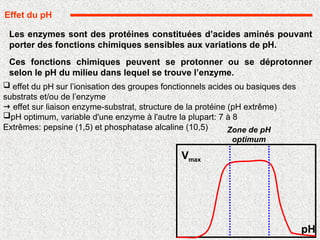
Les enzymes sontdes protéines constituées d’acides aminés pouvant
porter des fonctions chimiques sensibles aux variations de pH.
Ces fonctions chimiques peuvent se protonner ou se déprotonner
selon le pH du milieu dans lequel se trouve l’enzyme.
Effet du pH
pH
Vmax
Zone de pH
optimum
effet du pH sur l’ionisation des groupes fonctionnels acides ou basiques des
substrats et/ou de l’enzyme
effet sur liaison enzyme-substrat, structure de la protéine (pH extrême)
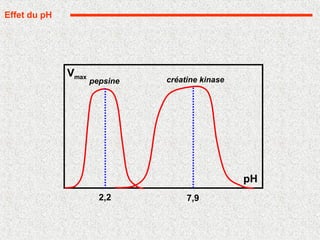
pH optimum, variable d'une enzyme à l'autre la plupart: 7 à 8
Extrêmes: pepsine (1,5) et phosphatase alcaline (10,5)
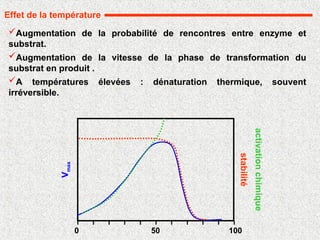
Augmentation de laprobabilité de rencontres entre enzyme et
substrat.
Augmentation de la vitesse de la phase de transformation du
substrat en produit .
A températures élevées : dénaturation thermique, souvent
irréversible.
Effet de la température
V
max
0 50
stabilité
activation
chimique
100
27.
Unités de vitesse: système international
vi = 600 µmoles de P / min
Par convention, il a été décidé que chaque fois qu’une quantité
d’enzyme donnée produirait 1 µmole de produit par min cela
correspondrait à 1 U ou 1 UIE (unité internationale enzymatique).
Exemple
vi = 600 U = 600 UIE
correspond à
Unités utilisées
28.
Unités de vitesse: système international
vi = 600 µmoles de P / min
Aujourd'hui, l'unité recommandée est le katal (kat). Elle correspond
à l’apparition d’une mole de produit par seconde (activité énorme
jamais retrouvée physiologiquement). On utilise donc des sous
multiples le mkat, le µkat, le nkat, etc…
Exemple
vi = 10 µmoles de P / sec
correspond à
donc à
vi = 10 µkat
Unités utilisées
5.6. Inhibiteurs
• Moléculemodifiant la réaction enzymatique en
influençant la liaison enzyme-substrat
• substances naturelles : modulation du métabolisme
– médicaments et agents toxiques
– outils expérimentaux (mécanisme enzymatique, voies métaboliques)
• Inhibition irréversible: souvent liaison covalente E-I stable
– diminution de la concentration de l’enzyme actif; ex l’aspirine inhibe
irréversiblement la prostaglandine synthétase .
• Inhibition réversible : liaison E-I non covalente
– inhibition compétitive
– inhibition non compétitive
– inhibition incompétitive
31.
E + P
ES
k1
k-1
k2
E+ S
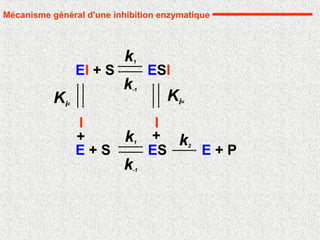
Mécanisme général d'une inhibition enzymatique
I
EI + S
+
k1
k-1
ESI
I
+
Kiu
Kic
32.
Inhibition compétitive
Il s'agitd'un exemple d'inhibition où le substrat et l'inhibiteur sont en
compétition pour le même site de fixation
On définit Kic
comme la constante de dissociation du complexe EI
k-1
E + P
ES
k1 k2
E + S
I
EI
+
Kic
complexe en "cul de sac"
I se fixe à E
33.
A. Inhibition compétitive
L’inhibiteuret le substrat possèdent une analogie
structural qui est connue par le site de fixation sur
l’enzyme
L’inhibiteur ne peut se combiner avec l’enzyme
en même temps que le substrat.
En présence d’inhibiteur, L’affinité diminue (Km
augmente)
Vmax reste inchangée
E + P
ES
k1
k-1
k2
E+ S
I
EI + S
+
k1
k-1
ESI
I
+
Kiu
Kic avec Kic
= Kiu
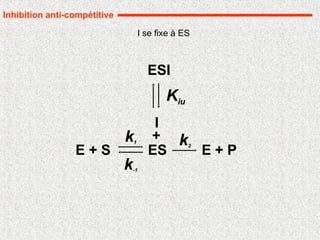
Inhibition non compétitive pure
I se fixe à E et ES avec la même affinité
37.
B. Inhibition noncompétitive
L’inhibiteur se combine avec l’enzyme
indépendamment du substrat : les deux peuvent
donc se fixer simultanément.
I bloque l’activité de l’enzyme
La fixation de I sur l’enzyme ne modifie pas Km.
l’inhibiteur se manifeste par la diminution de la
vitesse maximale
Cas particulier :inhibition irréversible
1
[S]
vi
1
[I]
1
Vmax
KM
1
-
E + P
ES
k1
k-1
k2
E + S
EI
I
+
k3
Cette représentation ressemble à celle d'une inhibition non-compétitive pure.
42.
état T (tendu)
conformationpassive
ligands: inhibiteurs
état R (relaché)
conformation active
ligands: substrats, stimulateurs
Allostérie
• enzymes constitués de plusieurs sous-unités plusieurs sites
actifs
Il existe au moins un site de fixation pour le substrat (site
actif, catalytique) et au moins un site de fixation (spécifique) pour
un modulateur (site régulateur).
• Les enzymes allostériques font intervenir une liaison réversible, non
covalente, d’une molécule régulatrice appelée modulateur, ou
effecteur, allostérique.
43.
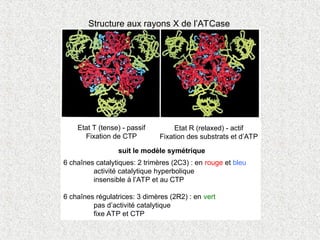
Structure aux rayonsX de l’ATCase
Etat T (tense) - passif
Fixation de CTP
Etat R (relaxed) - actif
Fixation des substrats et d’ATP
6 chaînes catalytiques: 2 trimères (2C3) : en rouge et bleu
activité catalytique hyperbolique
insensible à l’ATP et au CTP
6 chaînes régulatrices: 3 dimères (2R2) : en vert
pas d’activité catalytique
fixe ATP et CTP
suit le modèle symétrique
44.
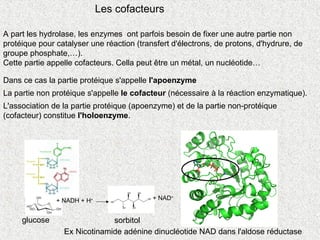
A part leshydrolase, les enzymes ont parfois besoin de fixer une autre partie non
protéique pour catalyser une réaction (transfert d'électrons, de protons, d'hydrure, de
groupe phosphate,…).
Cette partie appelle cofacteurs. Cella peut être un métal, un nucléotide…
Dans ce cas la partie protéique s'appelle l'apoenzyme
La partie non protéique s'appelle le cofacteur (nécessaire à la réaction enzymatique).
L'association de la partie protéique (apoenzyme) et de la partie non-protéique
(cofacteur) constitue l'holoenzyme.
Les cofacteurs
Ex Nicotinamide adénine dinucléotide NAD dans l'aldose réductase
+ NADH + H+ + NAD+
glucose sorbitol
Pour catalyser correctementune réaction chimique, un enzyme a
parfois besoin d'une molécule "exogène".
Il peut s’agir d’un ion ou d’un coenzyme.
Il y a deux familles de coenzymes :
- ceux qui ne sont que transitoirement liés à l’enzyme et que
l’on considère alors comme des co-substrats,
- ceux que l’on nomme groupes prosthétiques et qui sont
liés de façon covalente à la protéine.
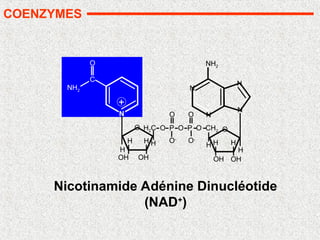
COENZYMES
Le cofacteur d’un enzyme est une molécule qui apporte un
groupement chimique important pour la catalyse enzymatique,
groupement que ne possède pas l’enzyme seul.
47.
Ion essentiel :2 types
• ions activateurs (lâchement liés)
– permet la liaison entre le substrat et les enzymes activées par les
métaux
– Ex: kinase et Mg++
• ions métalliques (fermement liés)
– rôle catalytique lors de sa liaison à la protéine métallo-enzymes
– Ex: la phosphotriesterase et Zn++
Le site actif de la phosphotriesterase est constitué de 2 atomes de zinc
48.
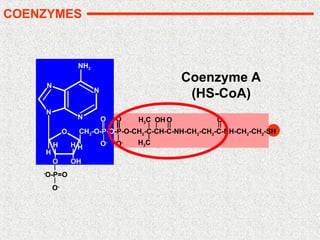
Coenzyme (molécule organique): 2 types
• cosubstrat (fixation transitoire à la protéine)
– subit une transformation au cours de la réaction
– Ex: NAD(P)/NAD(P)H : réactions d'oxydo-réduction
• groupe prosthétique (fixation permanente à la protéine)
– retrouve sa forme initiale en fin de réaction
– Ex: pyridoxal-5-phosphate : transfert de groupements aminés, biotine :
réaction de carboxylation
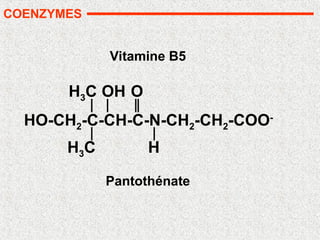
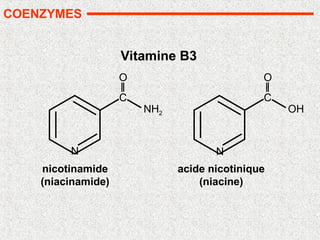
• Nombreuses vitamines: précurseurs de coenzymes
• Nicotinamide: NAD et NADP (B3)
• pyridoxine (B6)
• biotine (B8)
• acide folique (B9)
• cobalamine (B12)
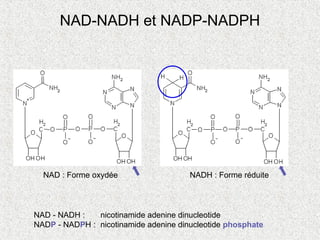
NAD-NADH et NADP-NADPH
NAD- NADH : nicotinamide adenine dinucleotide
NADP - NADPH : nicotinamide adenine dinucleotide phosphate
NAD : Forme oxydée NADH : Forme réduite
51.
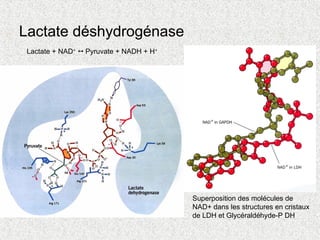
Lactate + NAD+
Pyruvate + NADH + H+
Superposition des molécules de
NAD+ dans les structures en cristaux
de LDH et Glycéraldéhyde-P DH
Lactate déshydrogénase
52.
COENZYMES
Biotine Carboxylation
Coenzyme àcobalamine
(vit B12)
Alkylation
Coenzyme A Transfert de groupe acyl
Acide folique Transfert de groupe acyl
Coenzymes à nicotinamide Oxydo-réduction
Phosphate de pyridoxal Transfert de groupe amino
Cofacteur Réaction impliquée
Coenzymes flaviniques Oxydo-réduction
Tétrahydrofolate Transfert de groupe à un carbone
Thiamine pyrophosphate Transfert de groupe aldéhyde

![milieu réactionnel
contenant à t=0 une
concentration de
substrat [S]0
Complexe enzyme-substrat](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-2-320.jpg)
![ajout de
l’enzyme [E]T
on est au temps 0
[E]T pour [E]TOTALE
milieu réactionnel
contenant à t=0 une
concentration de
substrat [S]0
Complexe enzyme-substrat](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-4-320.jpg)
![à t=0, la concentration en substrat est désignée [S]0
à t=0, la concentration en produit est égale à 0
la concentration en enzyme est désignée [E]T et reste
constante tout au long de la mesure
à t=0, la concentration du complexe enzyme-substrat, [ES],
est égale à 0
E + S E + P
ES
Complexe enzyme-substrat](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-6-320.jpg)
![Temps
Concentrations
[S]
[P]
[E]libre
[ES]
état pré-
stationnaire
état
stationnaire
état post-
stationnaire
[S]0
[E]libre
[P]
[ES]
Complexe enzyme-substrat
E + S E + P
ES](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-7-320.jpg)

![concentration en substrat [S]
vitesse
initiale
(Vi)
Vmax
[E]T cste
Équation d’une
hyperbole
Phases de la réaction](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-10-320.jpg)
![E + S E + P
ES
k1
k-1
k2
vi = k2 [ES]
La vitesse d’apparition du produit P (vitesse de la
réaction catalysée par l’enzyme) dépend de k2 et de la
concentration en ES.
La [ES] dépend de sa vitesse de formation et de sa
vitesse de disparition.
vformation ES = k1 [E] [S]
vdisparition ES = k-1 [ES] + k2 [ES] = (k-1 + k2) [ES]
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-11-320.jpg)
![Temps
Concentrations
[S]
[P]
[E]libre
[ES]
état pré-
stationnaire
état
stationnaire
état post-
stationnaire
E + S E + P
ES
[S]
[P]
[E]libre
[ES]
d[ES]
dt
=0
Vi
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-12-320.jpg)
![(k-1 + k2) [ES] = k1 [E] [S]
vformation ES = vdisparition ES
Pendant l'état dit "stationnaire" ou en "vitesse initiale", les
vitesses de formation et de disparition de ES sont égales.
Si on réarrange l’expression mathématique :
[ES] =
k1 [E] [S]
(k-1 + k2)
ou
[ES] =
[E] [S]
(k-1 + k2)
k1
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-13-320.jpg)
![KM =
(k-1 + k2)
k1
On simplifie l’équation en définissant une nouvelle constante, KM,
appelée constante de Michaelis et Menten :
Si on réarrange l’expression mathématique :
[ES] =
[E] [S]
KM
Équation de Michaelis-Menten
[ES] =
[E] [S]
(k-1 + k2)
k1](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-14-320.jpg)
![la [S] est égale à celle de départ ([S]0), puisque la concentration en
substrat est très largement supérieure à celle en enzyme.
[ES] =
[E] [S]
KM
La concentration en enzyme libre [E] doit être égale à la concentration
totale en enzyme ([E]T) moins la concentration en enzyme lié ([ES]).
[E] = [E]T – [ES]
[ES] =
[E] [S]
KM
devient [ES] =
([E]T – [ES]) [S]
KM
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-15-320.jpg)
![[ES] =
([E]T – [ES]) [S]
KM
Si on développe
cela devient [ES] =
[E]T [S] – [ES] [S]
KM
[ES] + =
[E]T [S]
KM
[ES] [S]
KM
[ES] =
[S]
KM
[E]T
1 +
[S]
KM
[ES] ( 1 + ) =
[E]T [S]
KM
[S]
KM
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-16-320.jpg)
![[ES] =
[S]
KM
[E]T
1 +
[S]
KM
Or on sait que : Vi = k2 [ES]
Donc on peut écrire : Vi = k2
[S]
KM
[E]T
1 +
[S]
KM
On peut simplifier :
vi = k2
[S]
KM
[E]T
+
[S]
KM
KM
KM
= k2
[E]T
[S]
[S]
KM +
Équation de Michaelis-Menten](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-17-320.jpg)
![On sait que lorsque toutes les molécules d’enzyme sont saturées,
[ES] = [E]T et la vitesse de transformation du substrat en produit
est alors maximale :
Vmax = k2 [E]T
L’équation de Michaelis-Menten devient :
vi = Vmax
[S]
[S]
KM +
Vitesse
initiale
Équation de Michaelis-Menten
vi = k2
[E]T
[S]
[S]
KM +](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-18-320.jpg)
![vi = Vmax
[S]
[S]
KM +
Si vi = ½ Vmax
½ Vmax = Vmax
[S]
[S]
KM +
½ (KM + [S]) = [S]
½ KM = [S] - ½ [S]
½ KM = ½ [S]
KM = [S]
KM est inversement proportionnelle à l’affinité de l’enzyme. Plus
l’affinité est élevée, et moins il faudra de substrat pour que
l’enzyme fonctionne.
Constante de Michaelis-Menten
KM est la concentration en substrat
pour laquelle l’enzyme fonctionne à
la moitié de sa vitesse maximale](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-19-320.jpg)
![KM s’exprime en unité de concentration de substrat.
KM = [S] lorsque vi = ½ Vmax
mole/L
µmole/L
mg/L
g/L
etc…
Constante de Michaelis-Menten
Nomenclature :
M signifie Michaelis mais,
officiellement, il faudrait
écrire cette constante Km](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-20-320.jpg)
![Détermination de Km et Vmax:
Graphiquement on ne peut pas déterminer Km
et Vmax a partir de la courbe v= f [s]
On préfère placer les points expérimentaux sur
un diagramme ou ils s’alignent sur une droite.
Parmi les différentes transformation de
l’equation de michaelis réaliser dans ce but: la
représentation de lineweaver et burk (L.B)la
représentation en double inverse.
Consiste à porter les variation de 1/v en
fonction de 1/ [s]](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-21-320.jpg)







![1
[S]
vi
1
1
Vmax
[I]
1
KM
-
Inhibition compétitive](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-35-320.jpg)


![Inhibition non compétitive pure
1
[S]
vi
1
[I]
1
Vmax
KM
1
-](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-38-320.jpg)
![KM abaissé
Vmax abaissée
KM
Vmax
V0
[S]
+ I
Inhibition incompétitive](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-40-320.jpg)
![Cas particulier : inhibition irréversible
1
[S]
vi
1
[I]
1
Vmax
KM
1
-
E + P
ES
k1
k-1
k2
E + S
EI
I
+
k3
Cette représentation ressemble à celle d'une inhibition non-compétitive pure.](https://image.slidesharecdn.com/enzymologie2-250226194006-3862a0c1/85/Enzymologie2-ppt-41-320.jpg)