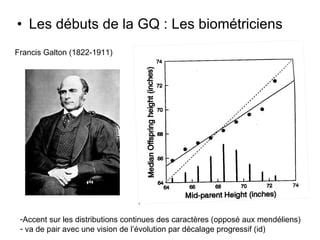
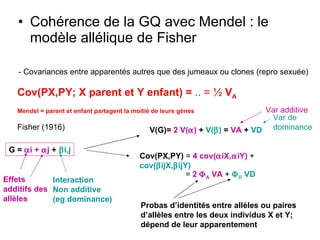
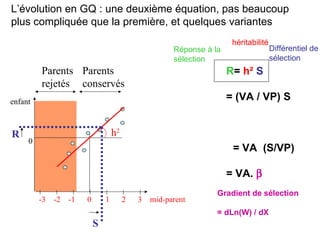
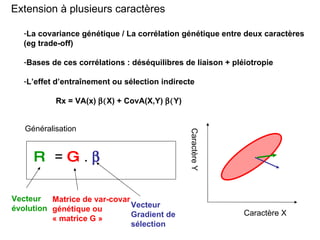
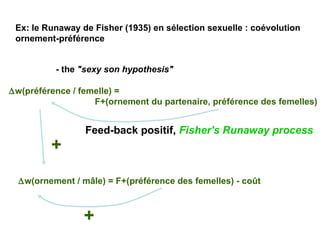
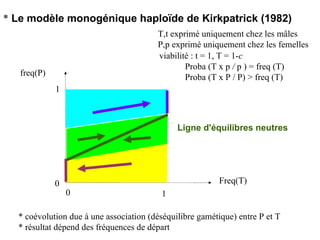
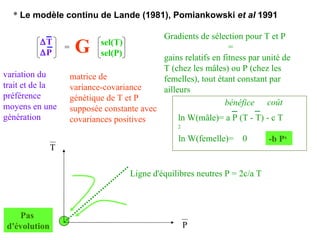
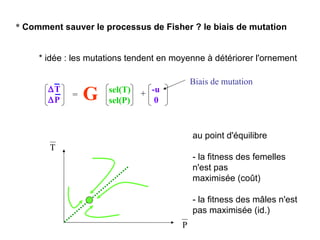
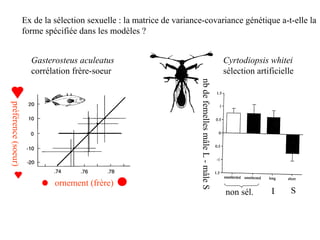
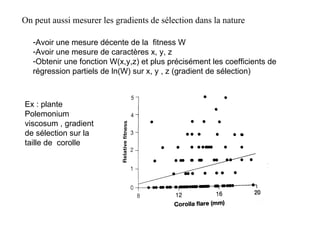
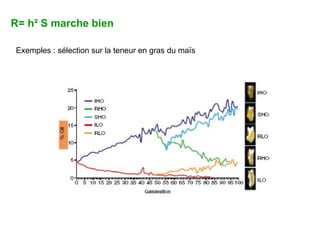
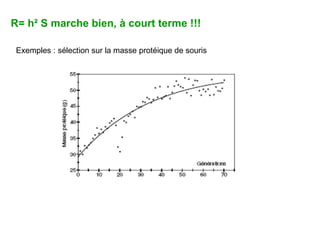
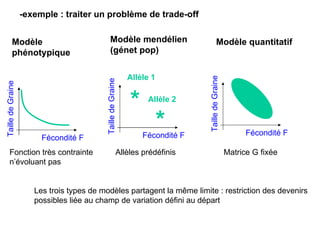
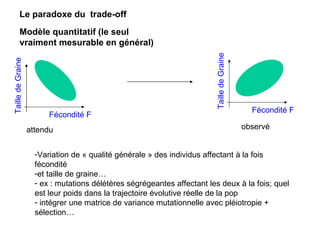
Le document présente la modélisation de l'évolution et de la génétique quantitative, en abordant trois méthodes principales : l'optimisation phénotypique, la théorie des jeux et la dynamique adaptative. Il décrit également les modèles d'hérédité, la transmission des caractères phénotypiques, et les limites de la génétique quantitative à long terme, tout en soulignant l'importance des covariances génétiques et de la mesure des variances. Enfin, il illustre ces concepts à travers des exemples, comme la sélection sexuelle et les modèles de Fisher, tout en mettant en avant les défis de la mesure en biologie évolutive.