Téléchargé 11 fois












![l’artefact d’A ttraction des L ongues B ranches [ Felsenstein (1978) Syst Zool 27:401 ] Philippe et al. (2000) Proc. Royal Soc. Lond. B 267:1213.](https://image.slidesharecdn.com/confevolutionprecocegouy-23590/85/ConfEvolutionPrecoceGouy-19-320.jpg)


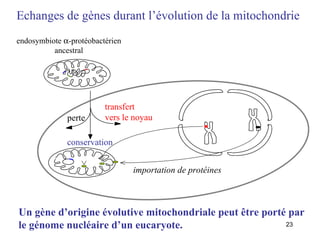
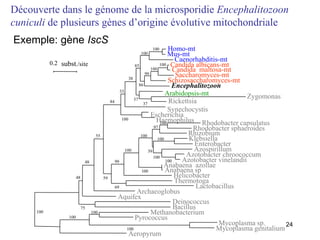
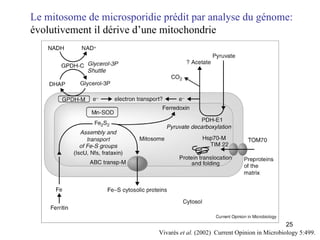
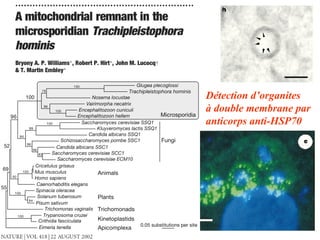

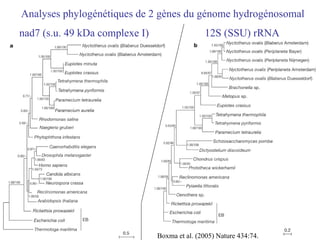
![Le chaînon manquant entre mitochondrie et hydrogénosome Fragment de 14027 pb du génome hydrogénosomal Découverte [Akhmanova et al. (1998) Nature 396:527] puis séquençage partiel du génome hydrogénosomal du cilié Nyctothermus ovalis Plusieurs protéines codées par le génome hydrogénosomal se groupent avec leurs homologues mitochondriaux de ciliés aérobies. Identification de plusieurs gènes nucléaires pour des composants du protéome mitochondrial (pyruvate déshydrogénase, complexe II).](https://image.slidesharecdn.com/confevolutionprecocegouy-23590/85/ConfEvolutionPrecoceGouy-29-320.jpg)
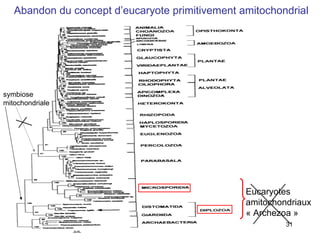
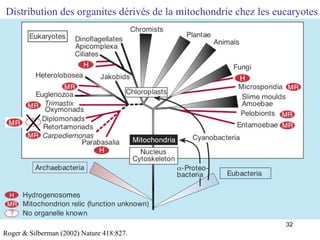


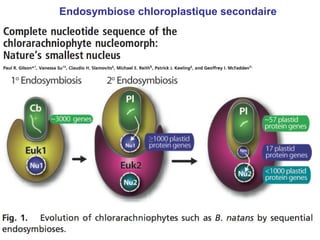
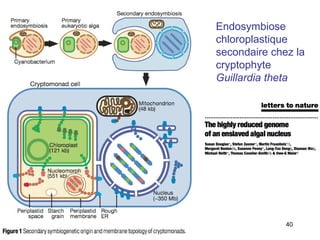
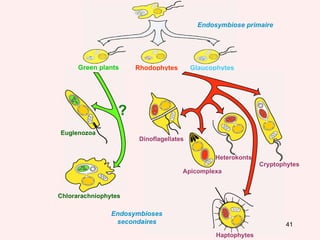
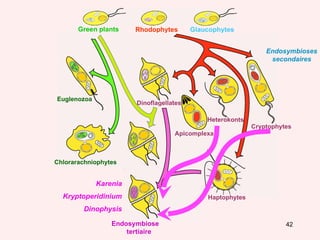

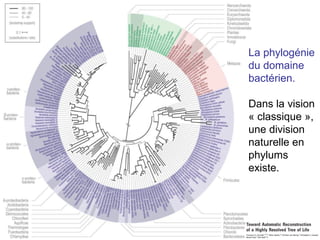

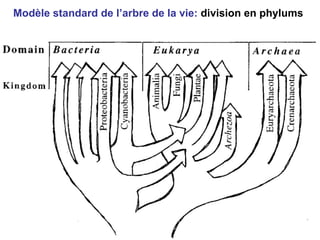
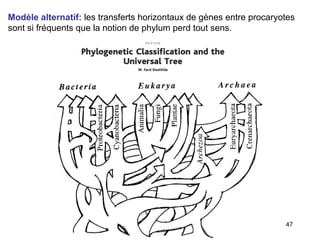
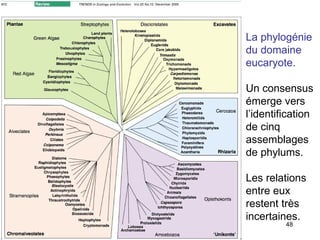

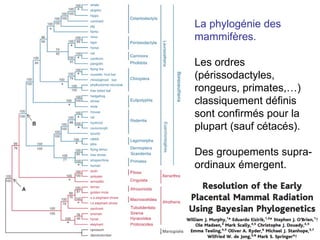

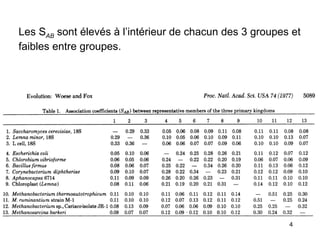
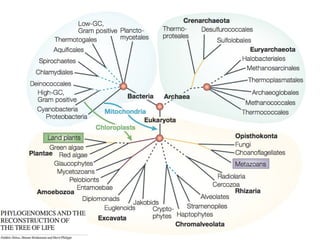
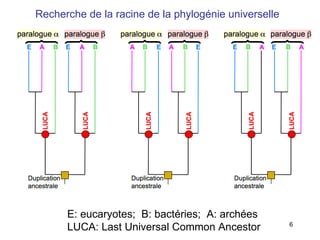
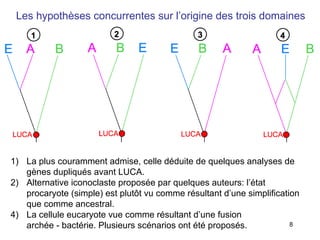
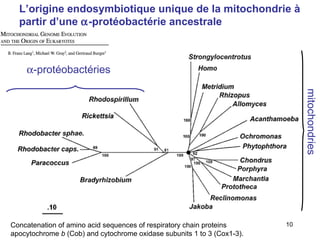
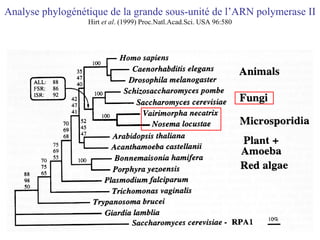
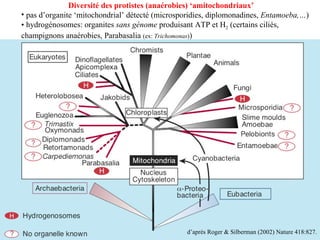
Le document traite des récentes découvertes concernant l'évolution des phases précoces de la vie, avec un accent sur l'origine des eucaryotes et la mitochondrie, ainsi que sur les hypothèses concurrentes de l'endosymbiose. Il explore également la diversité des eucaryotes amitochondriaux et les implications phylogénétiques des analyses moléculaires, tout en discutant des relations évolutives au sein des trois domaines de la vie. Enfin, il remet en question la simplicité du modèle traditionnel de l'arbre de la vie en faveur d'un modèle plus complexe incluant des transferts horizontaux de gènes.











































![Biologie cellulaire1[1].pptx...............](https://cdn.slidesharecdn.com/ss_thumbnails/biologiecellulaire11-251218173415-db5e4bf0-thumbnail.jpg?width=640&height=640&fit=bounds)












































