Téléchargé 56 fois















































































































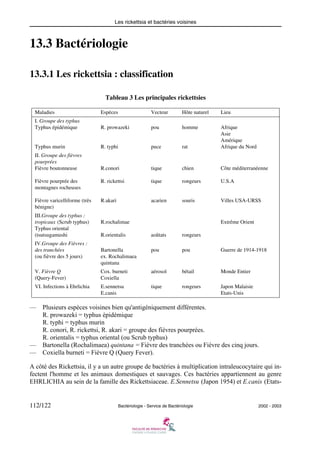











Ce document est un cours de bactériologie de l'Université Pierre et Marie Curie pour le niveau DCEM1, couvrant divers aspects des bactéries, y compris leur anatomie, génétique, et pathogénicité. Il détaille également l'étude des principales familles de bactéries, leur diagnostic et les traitements associés. La mise à jour date du 24 mars 2003 et le document comprend des chapitres et une table des matières exhaustive.



































































